

Disruption of a rice pentatricopeptide repeat protein causes a seedling-specific albino phenotype and its utilization to enhance seed purity in hybrid rice production
- PMID: 22430843
- PMCID: PMC3366715
- DOI: 10.1104/pp.112.195081
Disruption of a rice pentatricopeptide repeat protein causes a seedling-specific albino phenotype and its utilization to enhance seed purity in hybrid rice production
Abstract
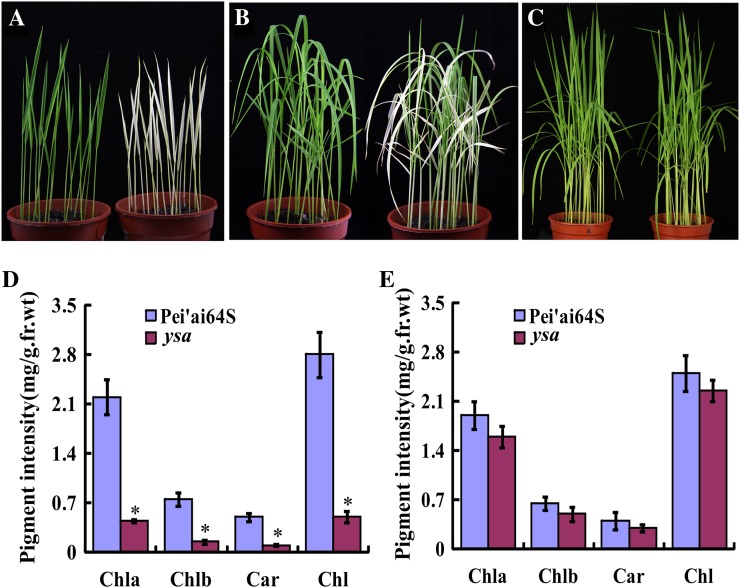
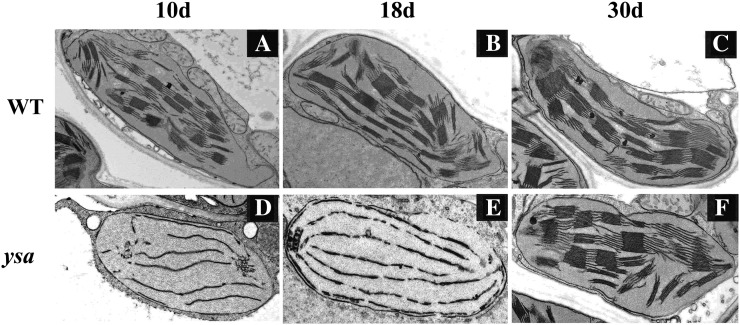
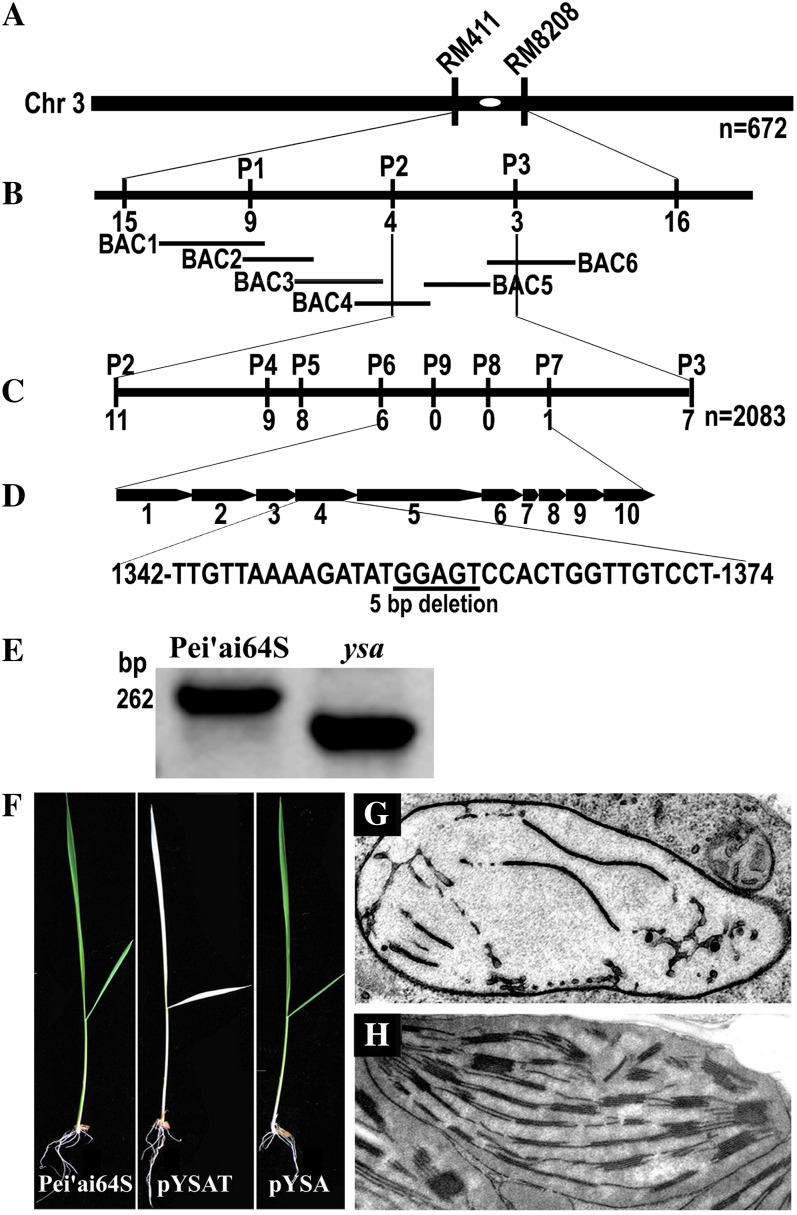
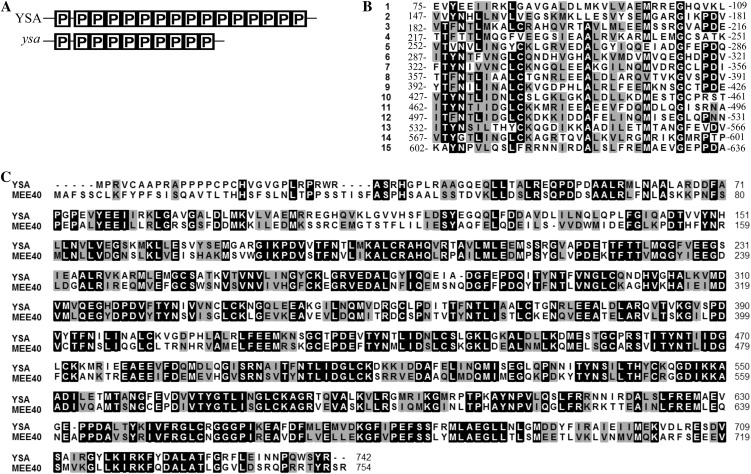
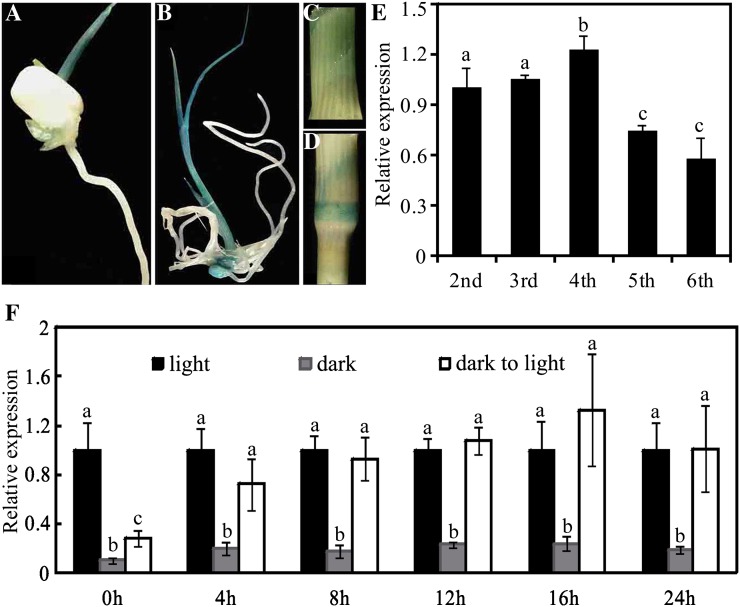
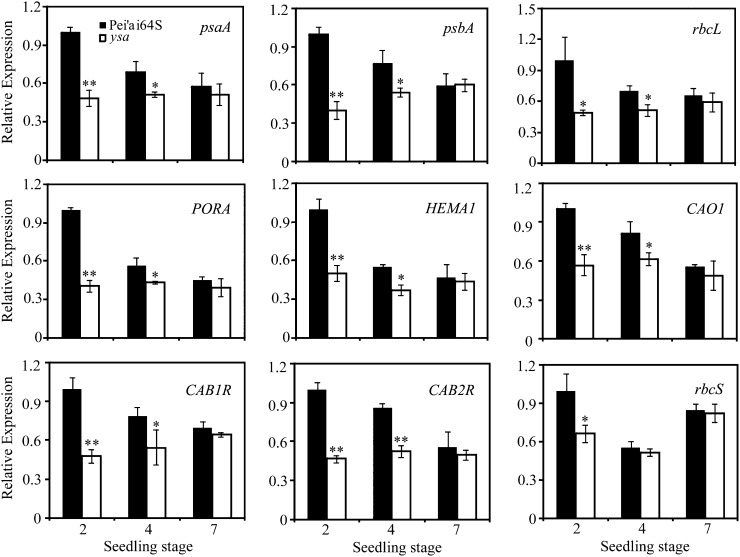
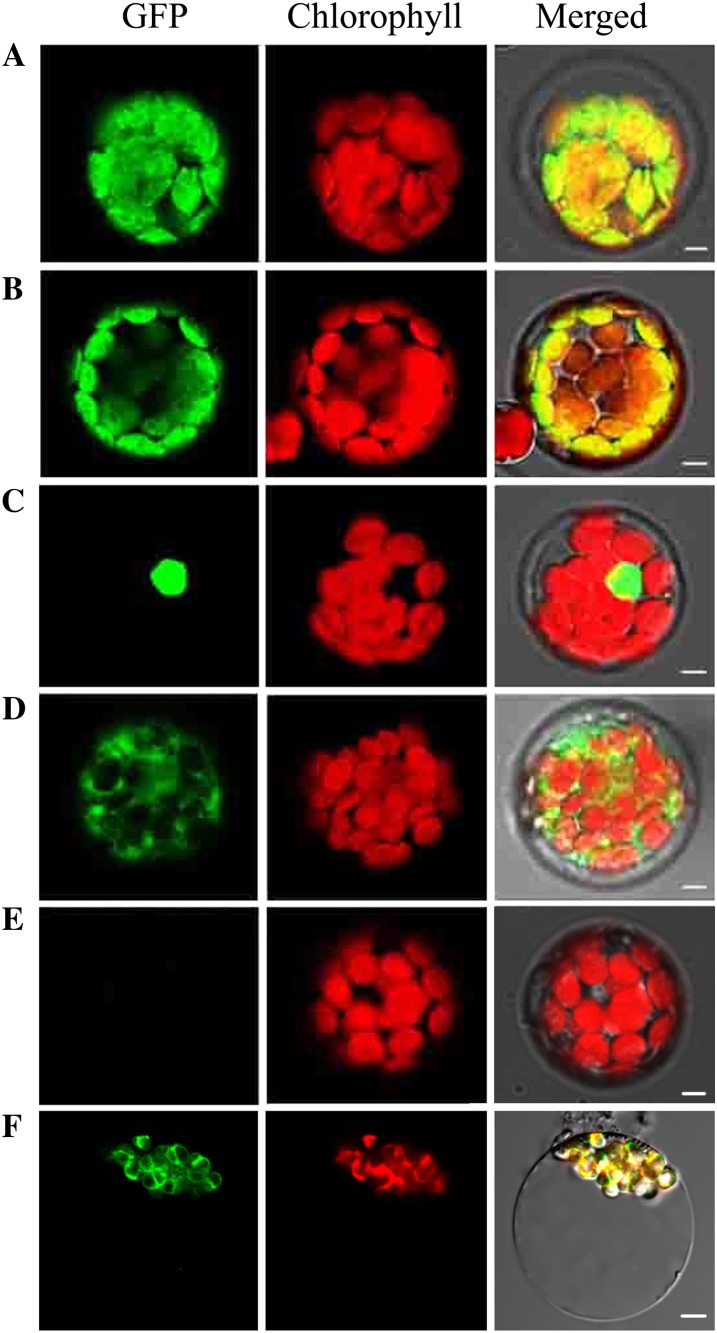
The pentatricopeptide repeat (PPR) gene family represents one of the largest gene families in higher plants. Accumulating data suggest that PPR proteins play a central and broad role in modulating the expression of organellar genes in plants. Here we report a rice (Oryza sativa) mutant named young seedling albino (ysa) derived from the rice thermo/photoperiod-sensitive genic male-sterile line Pei'ai64S, which is a leading male-sterile line for commercial two-line hybrid rice production. The ysa mutant develops albino leaves before the three-leaf stage, but the mutant gradually turns green and recovers to normal green at the six-leaf stage. Further investigation showed that the change in leaf color in ysa mutant is associated with changes in chlorophyll content and chloroplast development. Map-based cloning revealed that YSA encodes a PPR protein with 16 tandem PPR motifs. YSA is highly expressed in young leaves and stems, and its expression level is regulated by light. We showed that the ysa mutation has no apparent negative effects on several important agronomic traits, such as fertility, stigma extrusion rate, selfed seed-setting rate, hybrid seed-setting rate, and yield heterosis under normal growth conditions. We further demonstrated that ysa can be used as an early marker for efficient identification and elimination of false hybrids in commercial hybrid rice production, resulting in yield increases by up to approximately 537 kg ha(-1).
Figures

References
-
- Ai PH, Sun SB, Zhao JN, Fan XR, Xin WJ, Guo Q, Yu L, Shen QR, Wu P, Miller AJ, et al. (2009) Two rice phosphate transporters, OsPht1;2 and OsPht1;6, have different functions and kinetic properties in uptake and translocation. Plant J 57: 798–809 - PubMed
-
- Arabidopsis Genome Initiative (2000) Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408: 796–815 - PubMed
-
- Bi C, Li Z, Wan J. (1990) Effect of the lower temperature in mid-summer on the fertility stability of HPGMR. Chin J Rice Sci 4: 181–184
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
