

Adult-born hippocampal neurons promote cognitive flexibility in mice
- PMID: 22431384
- PMCID: PMC4784987
- DOI: 10.1002/hipo.22013
Adult-born hippocampal neurons promote cognitive flexibility in mice
Abstract
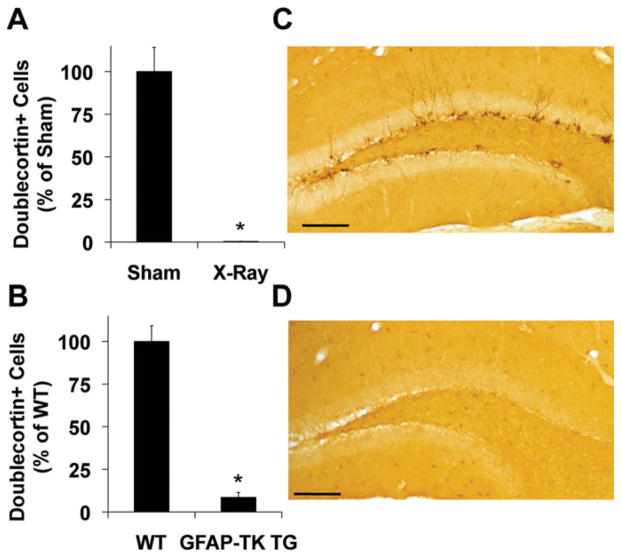
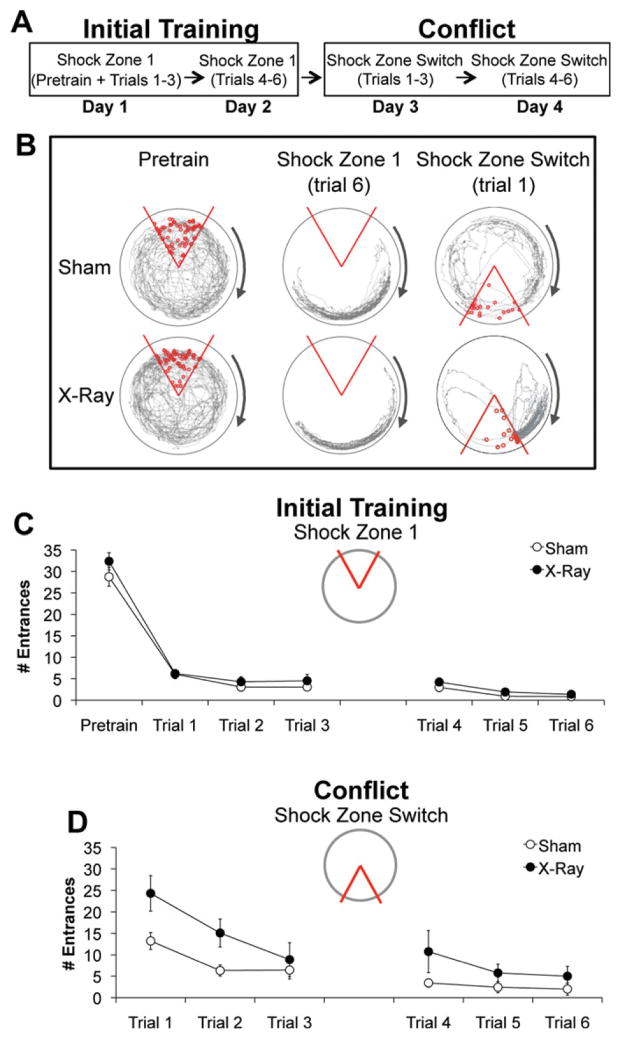
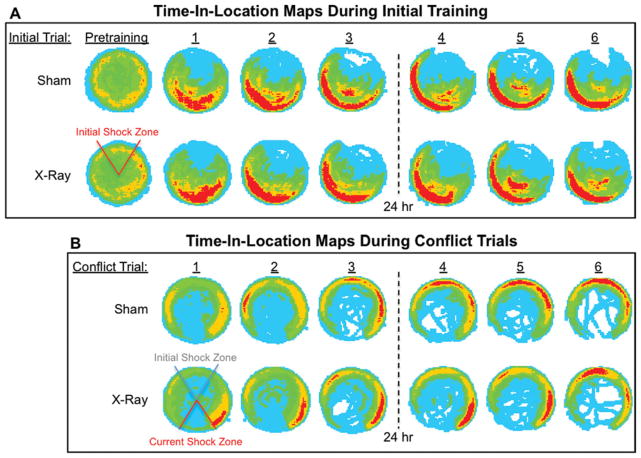
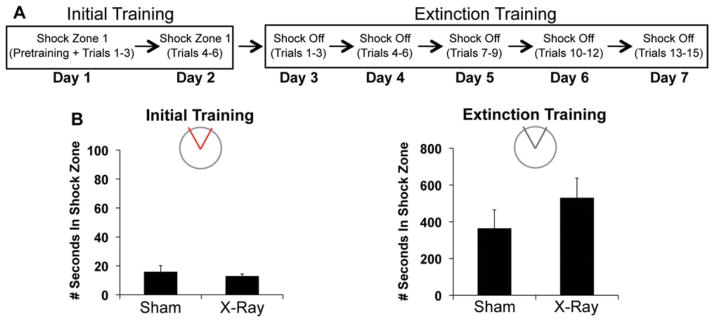
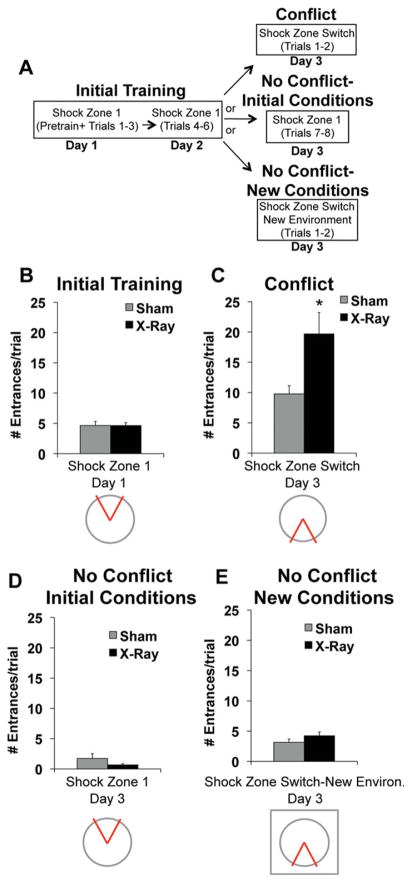
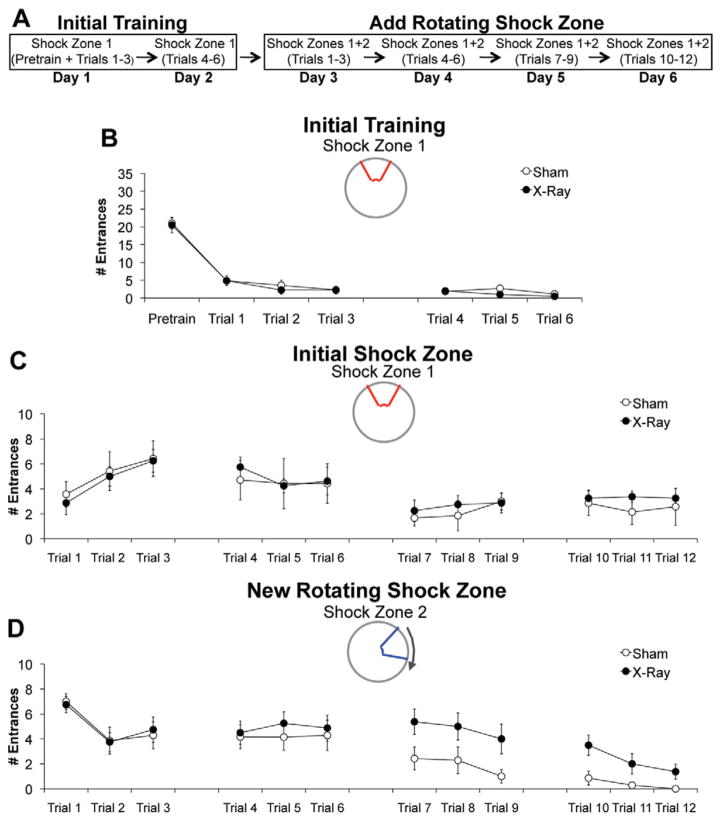
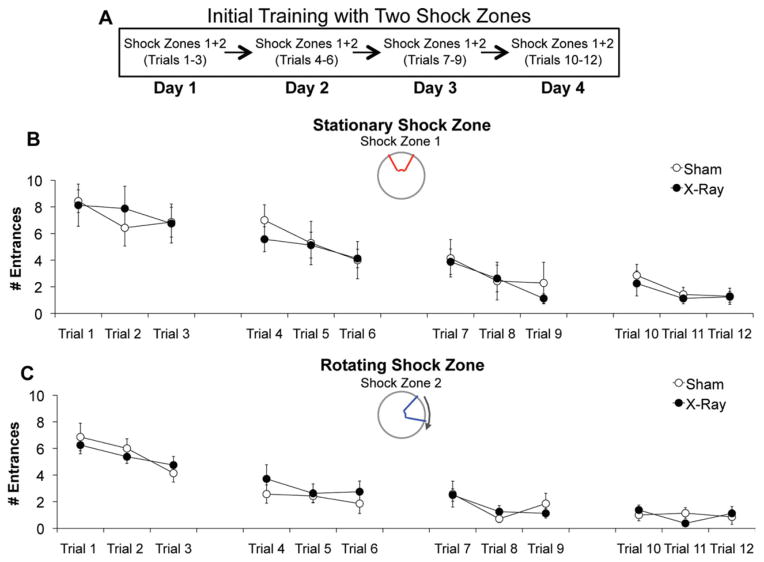
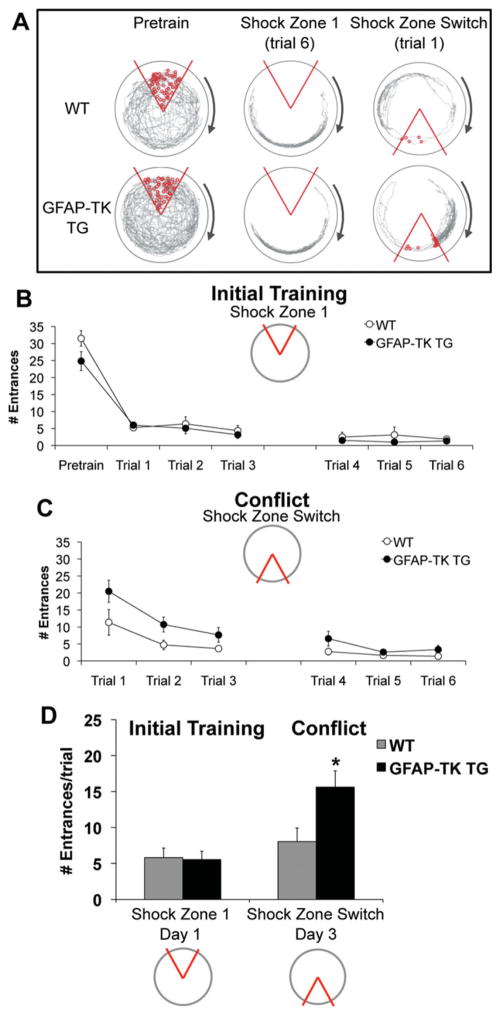
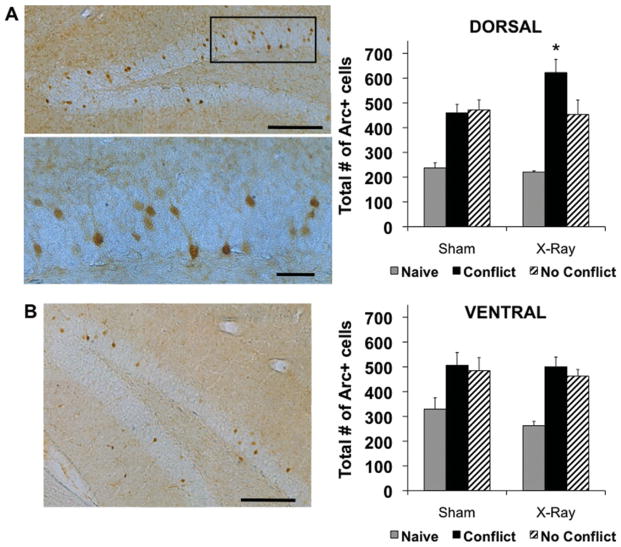
The hippocampus is involved in segregating memories, an ability that utilizes the neural process of pattern separation and allows for cognitive flexibility. We evaluated a proposed role for adult hippocampal neurogenesis in cognitive flexibility using variants of the active place avoidance task and two independent methods of ablating adult-born neurons, focal X-irradiation of the hippocampus, and genetic ablation of glial fibrillary acidic protein positive neural progenitor cells, in mice. We found that ablation of adult neurogenesis did not impair the ability to learn the initial location of a shock zone. However, when conflict was introduced by switching the location of the shock zone to the opposite side of the room, irradiated and transgenic mice entered the new shock zone location significantly more than their respective controls. This impairment was associated with increased upregulation of the immediate early gene Arc in the dorsal dentate gyrus, suggesting a role for adult neurogenesis in modulating network excitability and/or synaptic plasticity. Additional experiments revealed that irradiated mice were also impaired in learning to avoid a rotating shock zone when it was added to an initially learned stationary shock zone, but were unimpaired in learning the identical simultaneous task variant if it was their initial experience with place avoidance. Impaired avoidance could not be attributed to a deficit in extinction or an inability to learn a new shock zone location in a different environment. Together these results demonstrate that adult neurogenesis contributes to cognitive flexibility when it requires changing a learned response to a stimulus-evoked memory.
Copyright © 2012 Wiley Periodicals, Inc.
Figures

References
-
- Aimone JB, Wiles J, Gage FH. Potential role for adult neurogenesis in the encoding of time in new memories. Nat Neurosci. 2006;9:723–727. - PubMed
-
- Alvarez-Buylla A, Lim DA. For the long run: Maintaining germinal niches in the adult brain. Neuron. 2004;41:683–686. - PubMed
-
- Becker S. A computational principle for hippocampal learning and neurogenesis. Hippocampus. 2005;15:722–738. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
