

Review
doi: 10.1083/jcb.201109041.
The evolution of metazoan extracellular matrix
Affiliations
- PMID: 22431747
- PMCID: PMC3308698
- DOI: 10.1083/jcb.201109041
Item in Clipboard
Review
The evolution of metazoan extracellular matrix
J Cell Biol.
.
Abstract
The modular domain structure of extracellular matrix (ECM) proteins and their genes has allowed extensive exon/domain shuffling during evolution to generate hundreds of ECM proteins. Many of these arose early during metazoan evolution and have been highly conserved ever since. Others have undergone duplication and divergence during evolution, and novel combinations of domains have evolved to generate new ECM proteins, particularly in the vertebrate lineage. The recent sequencing of several genomes has revealed many details of this conservation and evolution of ECM proteins to serve diverse functions in metazoa.
Figures
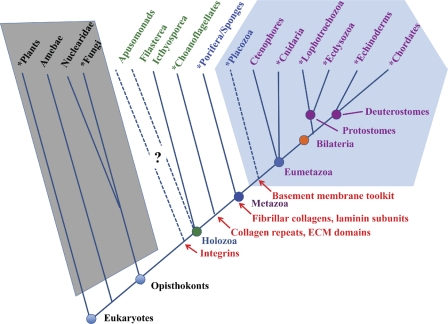
Eukaryotic phylogeny and the appearance of ECM proteins. The figure diagrams the major taxonomic divisions of eukaryotes (Box 1), and is based on published phylogenetic analyses (e.g., Philippe et al., 2005, 2009; Shalchian-Tabrizi et al., 2008; Pick et al., 2010) as well as on analyses of whole genome sequences (see main text). Taxa for which whole genome sequences are available are marked with asterisks. The diagram represents topologies of relationships, and the branch lengths are not intended to reflect accurate evolutionary distances. The relationships of apusomonads and placozoa are not well defined; their placement in the diagram is influenced in part by evidence from analyses of ECM-related genes (see main text). Taxa with the complement of ECM proteins typical of all bilateria are within the light blue hexagon. Taxa highlighted with green or blue text have some homologues of bilaterian ECM proteins or their receptors (see main text), but lack a complete set. The complexity of the known ECM protein sets in these organisms increases from left to right. In contrast, the taxa within the gray trapezoid show no evidence of any credible examples of metazoan ECM proteins. First known appearances of relevant genes/proteins or domains are marked in red. Note that taxa appearing to the right of (i.e., subsequent to) the origin of a given feature may have lost it; examples would be the absence of integrins in choanoflagellates sequenced to date and the absence of fibrillar collagens in Drosophila (in both cases, presumably by gene loss).
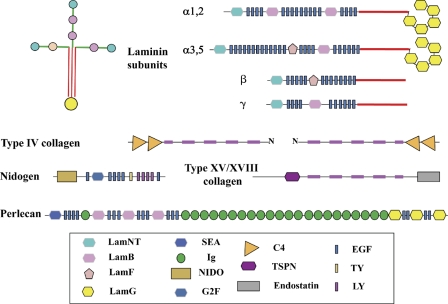
The basement membrane toolkit. The figure shows the domain structures of the core proteins of metazoan basement membranes, based on domain predictions largely using SMART and Pfam domain definitions. All bilaterian clades encode this set of nine proteins in their genomes. There are typically two distinct laminin α subunits and one each of the laminin β and γ subunits. Each subunit has a characteristic domain organization. A laminin protomer is an αβγ trimer (shown at the top left) associated through coiled-coil domains in each subunit (red) and disulfide bonds (not depicted). Type IV collagen is a trimer of two homologous subunits, α1 and α2, usually adjacent in the genome in a head-to-head arrangement with a single promoter between the two genes. Signature pairs of C4 domains lie at the C termini of all type IV collagens, and the collagen segment (fuschia) is interrupted, allowing flexibility. Type IV collagen protomers associate through their N and C termini and through disulfide bonding to form a “chicken wire” network that provides structural strength to the basement membrane. Laminins bind to the collagen network and to nidogen. Perlecan, which is a complex heparan sulfate proteoglycan, is also incorporated into the basement membrane. Two other collagens, types XV and XVIII, are also associated with vertebrate basement membranes, and an orthologue is present in all bilaterian clades. The high degree of conservation of this “toolkit” over more than half a billion years testifies to the essentiality of basement membranes.
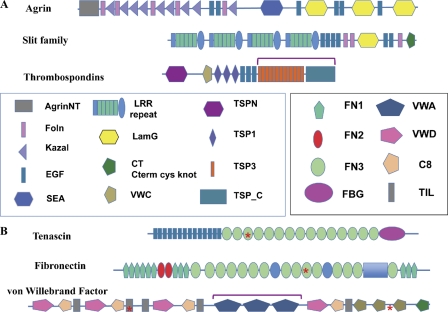
Examples of ECM proteins. (A) Proteins common to all bilaterian clades: agrin, slits, and thrombospondins. The figure shows domain maps of vertebrate proteins. Agrin is involved in synapse organization and slit family members in axonal guidance, although both are also involved in other processes as well. Proteins with similar arrangements of domains are found in all bilaterian clades, although occasionally missing domains, at least according to current gene predictions. Thrombospondins are characterized by the C-terminal set of domains (bracketed), which can be associated with a variety of additional N-terminal domains (Bentley and Adams, 2010; Adams and Lawler, 2011). The particular set of domains shown (including TSPN, VWC, and TSP1 repeats) is that of so-called type A thrombospondins. Type A thrombospondins are found only in deuterostomes, but all bilateria encode examples of type B thrombospondins, which lack the TSPN, VWC, and TSP1 domains but often have EGF repeats and sometimes others. (B) Proteins found only in the chordate lineage: tenascin, fibronectin, and VWF. These proteins evolved in the deuterostome lineage (see main text) and exemplify different sorts of domain shuffling. Tenascins are built from ancient domains (EGF, FN3, and FBG) found in many proteins throughout metazoa (and even in lower organisms). However, the particular set of domains in tenascins appears first in Branchiostoma (amphioxus), and the family is expanded in vertebrates. Fibronectins are built from a mixture of ancient domains (FN3), more recent ones (FN2), and chordate-specific domains (FN1). The only true fibronectins are vertebrate-specific and, within that subphylum, highly conserved and essential. VWF is based on an ancient gene structure (mucins) altered by insertion of three VWA domains (bracketed), which incorporate many of the key functions of VWF in hemostasis in vertebrates. All three of these characteristic vertebrate ECM proteins contain RGD motifs (asterisks), which are sites for binding of integrins.
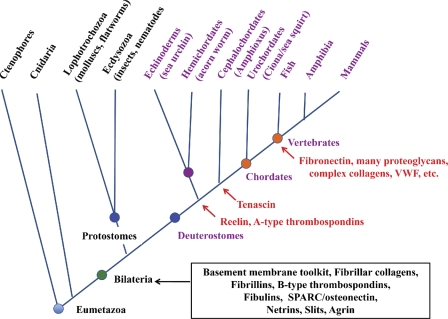
Deuterostome phylogeny and elaboration of ECM proteins. The figure diagrams deuterostome evolution according to the same principles outlined in Fig. 1. The core set of ECM proteins encoded in the genomes of all bilateria is boxed in black. Many of these proteins are also found in cnidaria (see main text). The main taxa of the deuterostome lineage and their relationships as currently understood are indicated with representative animals noted. The first reported appearances of particular proteins are marked in red.
References
-
- Aouacheria A., Geourjon C., Aghajari N., Navratil V., Deléage G., Lethias C., Exposito J.-Y. 2006. Insights into early extracellular matrix evolution: spongin short chain collagen-related proteins are homologous to basement membrane type IV collagens and form a novel family widely distributed in invertebrates. Mol. Biol. Evol. 23:2288–2302 10.1093/molbev/msl100 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
