

UU/UA dinucleotide frequency reduction in coding regions results in increased mRNA stability and protein expression
- PMID: 22434136
- PMCID: PMC3345983
- DOI: 10.1038/mt.2012.29
UU/UA dinucleotide frequency reduction in coding regions results in increased mRNA stability and protein expression
Abstract
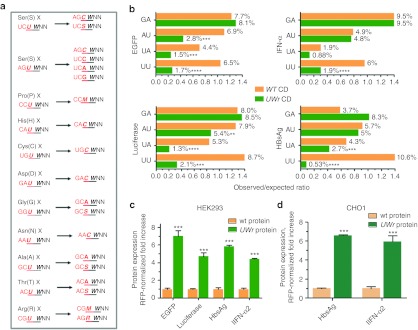
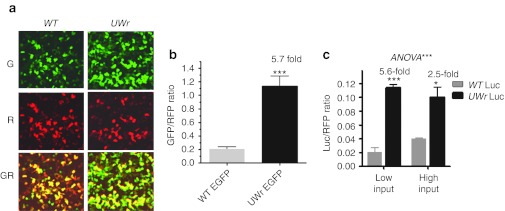
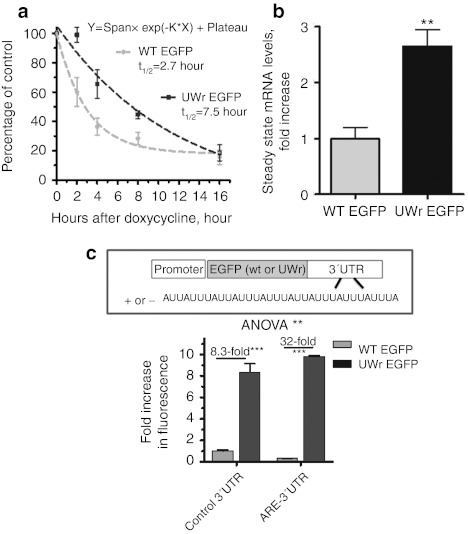
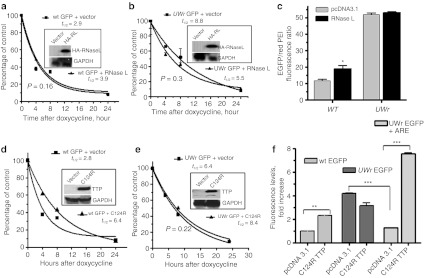
UU and UA dinucleotides are rare in mammalian genes and may offer natural selection against endoribonuclease-mediated mRNA decay. This study hypothesized that reducing UU and UA (UW) dinucleotides in the mRNA-coding sequence, including the codons and the dicodon boundaries, may promote resistance to mRNA decay, thereby increasing protein production. Indeed, protein expression from UW-reduced coding regions of enhanced green fluorescent protein (EGFP), luciferase, interferon-α, and hepatitis B surface antigen (HBsAg) was higher when compared to the wild-type protein expression. The steady-state level of UW-reduced EGFP mRNA was higher and the mRNA half-life was also longer. Ectopic expression of the endoribonuclease, RNase L, did not reduce the wild type or UW-reduced mRNA. A mutant form of the mRNA decay-promoting protein, tristetraprolin (TTP/ZFP36), which has a point mutation in the zinc-finger domain (C124R), was used. The wild-type EGFP mRNA but not the UW-reduced mRNA responded to the dominant negative action of the C124R ZFP36/TTP mutant. The results indicate the efficacy of the described rational approach to formulate a general scheme for boosting recombinant protein production in mammalian cells.
Figures
References
-
- Chekulaeva M., and, Filipowicz W. Mechanisms of miRNA-mediated post-transcriptional regulation in animal cells. Curr Opin Cell Biol. 2009;21:452–460. - PubMed
-
- Fritz DT, Bergman N, Kilpatrick WJ, Wilusz CJ., and, Wilusz J. Messenger RNA decay in mammalian cells: the exonuclease perspective. Cell Biochem Biophys. 2004;41:265–278. - PubMed
-
- Duan J., and, Antezana MA. Mammalian mutation pressure, synonymous codon choice, and mRNA degradation. J Mol Evol. 2003;57:694–701. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
