

Unusual combinatorial involvement of poly-A/T tracts in organizing genes and chromatin in Dictyostelium
- PMID: 22434426
- PMCID: PMC3371697
- DOI: 10.1101/gr.131649.111
Unusual combinatorial involvement of poly-A/T tracts in organizing genes and chromatin in Dictyostelium
Abstract
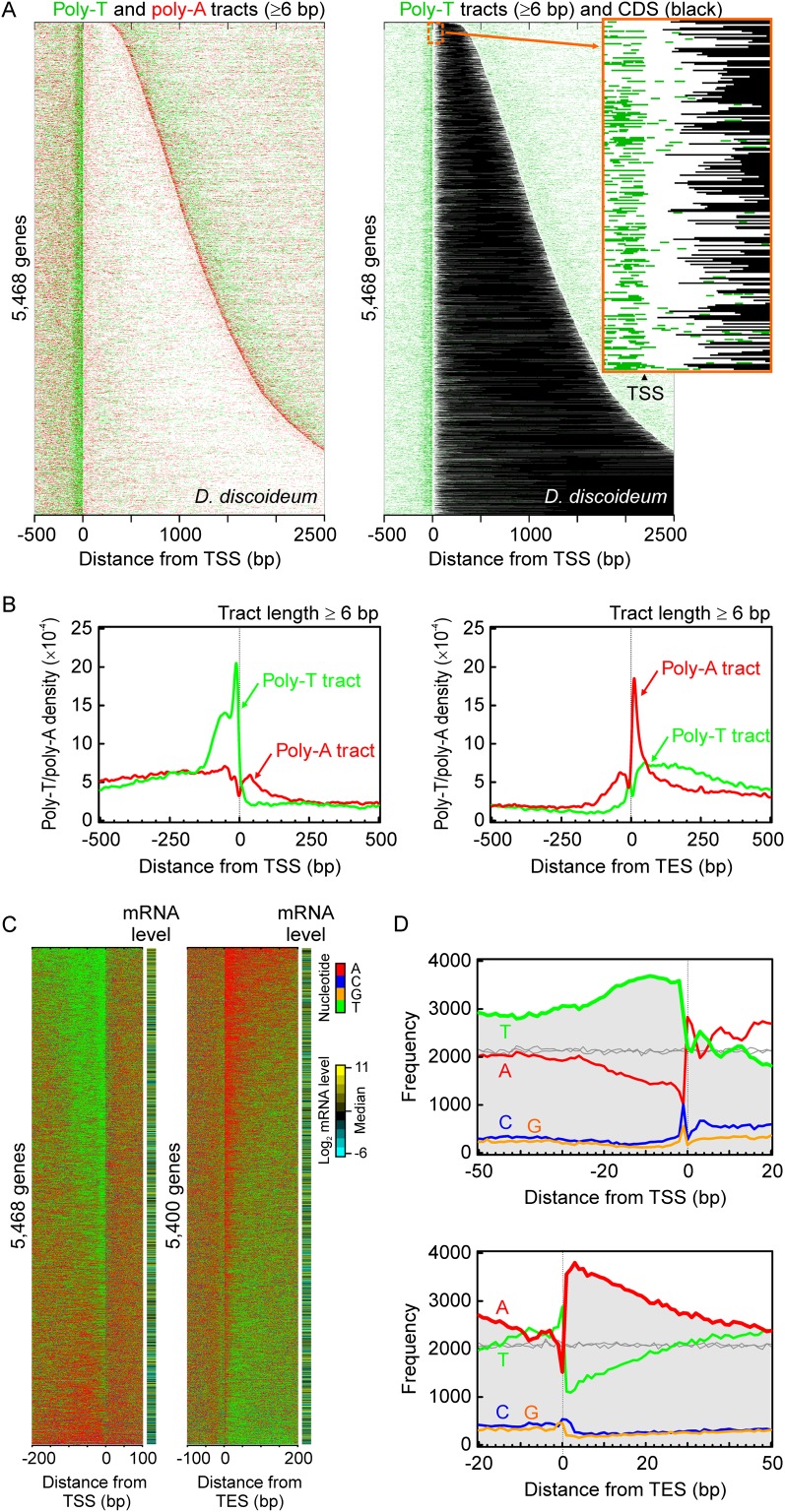
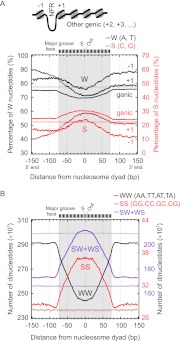
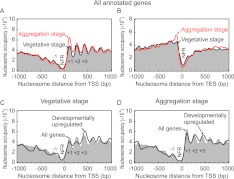
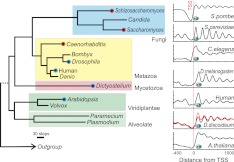
Dictyostelium discoideum is an amoebozoa that exists in both a free-living unicellular and a multicellular form. It is situated in a deep branch in the evolutionary tree and is particularly noteworthy in having a very A/T-rich genome. Dictyostelium provides an ideal system to examine the extreme to which nucleotide bias may be employed in organizing promoters, genes, and nucleosomes across a genome. We find that Dictyostelium genes are demarcated precisely at their 5' ends by poly-T tracts and precisely at their 3' ends by poly-A tracts. These tracts are also associated with nucleosome-free regions and are embedded with precisely positioned TATA boxes. Homo- and heteropolymeric tracts of A and T demarcate nucleosome border regions. Together, these findings reveal the presence of a variety of functionally distinct polymeric A/T elements. Strikingly, Dictyostelium chromatin may be organized in di-nucleosome units but is otherwise organized as in animals. This includes a +1 nucleosome in a position that predicts the presence of a paused RNA polymerase II. Indeed, we find a strong phylogenetic relationship between the presence of the NELF pausing factor and positioning of the +1 nucleosome. Pausing and +1 nucleosome positioning may have coevolved in animals.
Figures


References
-
- Albert I, Mavrich TN, Tomsho LP, Qi J, Zanton SJ, Schuster SC, Pugh BF 2007. Translational and rotational settings of H2A.Z nucleosomes across the Saccharomyces cerevisiae genome. Nature 446: 572–576 - PubMed
-
- Bailey TL, Elkan C 1994. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc Int Conf Intell Syst Mol Biol 2: 28–36 - PubMed
-
- Baldauf SL, Roger AJ, Wenk-Siefert I, Doolittle WF 2000. A kingdom-level phylogeny of eukaryotes based on combined protein data. Science 290: 972–977 - PubMed
-
- Basehoar AD, Zanton SJ, Pugh BF 2004. Identification and distinct regulation of yeast TATA box-containing genes. Cell 116: 699–709 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources