

Evaluation of pneumonia virus of mice as a possible human pathogen
- PMID: 22438539
- PMCID: PMC3347304
- DOI: 10.1128/JVI.00163-12
Evaluation of pneumonia virus of mice as a possible human pathogen
Abstract
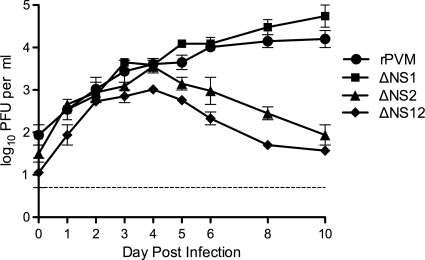
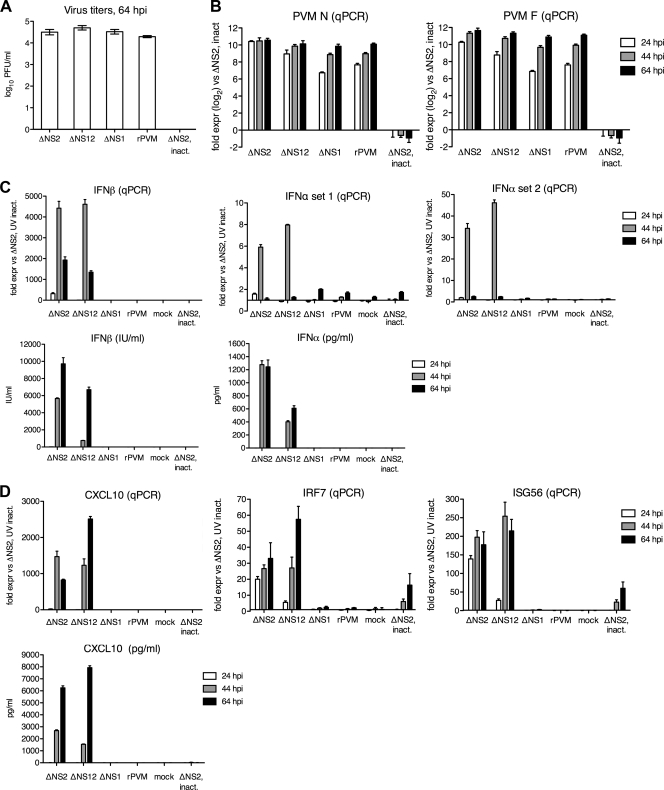
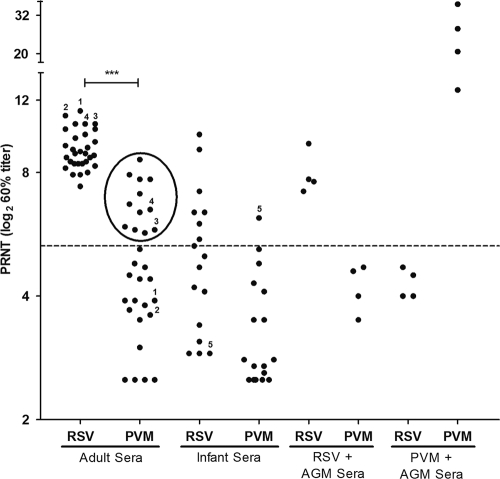
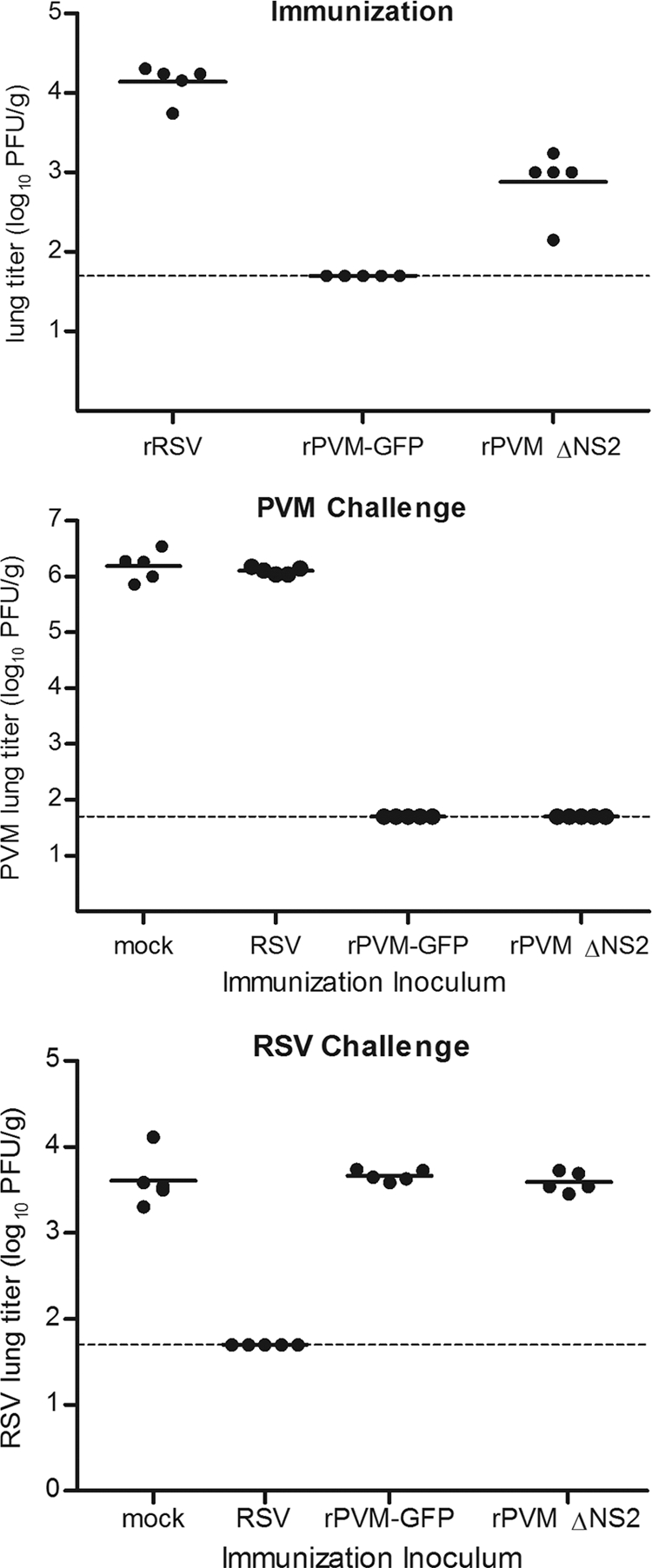
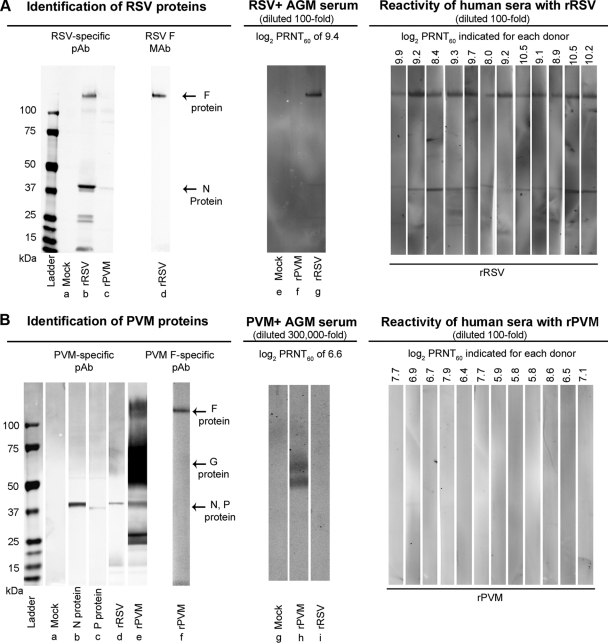
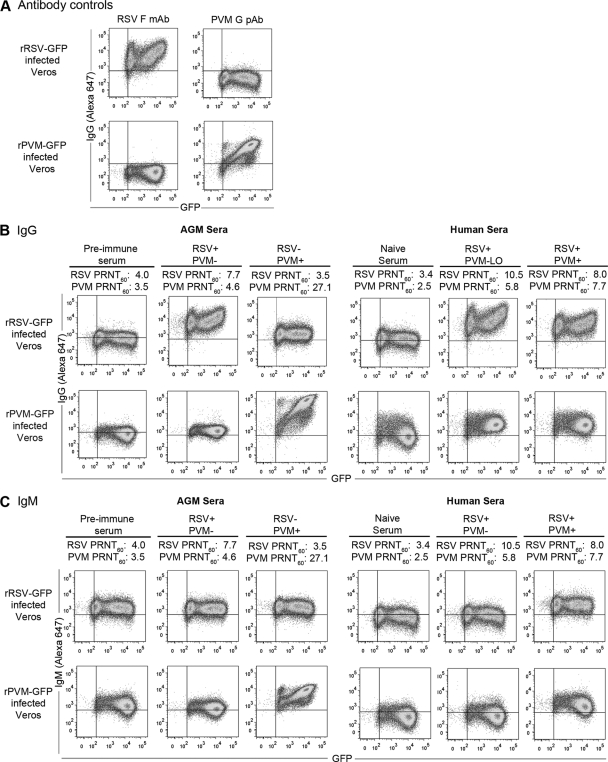
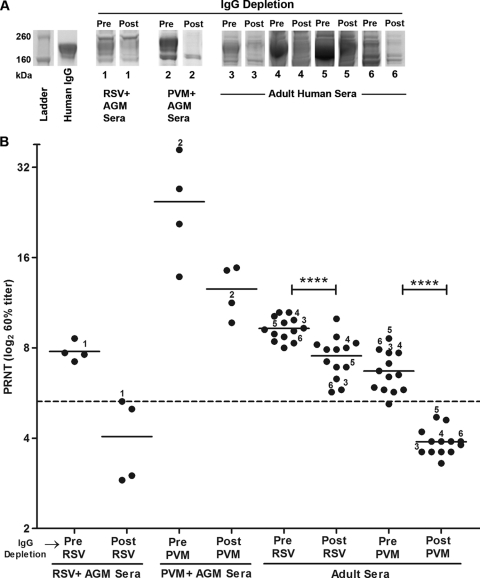
Pneumonia virus of mice (PVM), a relative of human respiratory syncytial virus (RSV), causes respiratory disease in mice. There is serologic evidence suggesting widespread exposure of humans to PVM. To investigate replication in primates, African green monkeys (AGM) and rhesus macaques (n = 4) were inoculated with PVM by the respiratory route. Virus was shed intermittently at low levels by a subset of animals, suggesting poor permissiveness. PVM efficiently replicated in cultured human cells and inhibited the type I interferon (IFN) response in these cells. This suggests that poor replication in nonhuman primates was not due to a general nonpermissiveness of primate cells or poor control of the IFN response. Seroprevalence in humans was examined by screening sera from 30 adults and 17 young children for PVM-neutralizing activity. Sera from a single child (6%) and 40% of adults had low neutralizing activity against PVM, which could be consistent with increasing incidence of exposure following early childhood. There was no cross-reaction of human or AGM sera between RSV and PVM and no cross-protection in the mouse model. In native Western blots, human sera reacted with RSV but not PVM proteins under conditions in which AGM immune sera reacted strongly. Serum reactivity was further evaluated by flow cytometry using unfixed Vero cells infected with PVM or RSV expressing green fluorescent protein (GFP) as a measure of viral gene expression. The reactivity of human sera against RSV-infected cells correlated with GFP expression, whereas reactivity against PVM-infected cells was low and uncorrelated with GFP expression. Thus, PVM specificity was not evident. Our results indicate that the PVM-neutralizing activity of human sera is not due to RSV- or PVM-specific antibodies but may be due to low-affinity, polyreactive natural antibodies of the IgG subclass. The absence of PVM-specific antibodies and restriction in nonhuman primates makes PVM unlikely to be a human pathogen.
Figures
References
-
- Anh DB, Faisca P, Desmecht DJ. 2006. Differential resistance/susceptibility patterns to pneumovirus infection among inbred mouse strains. Am. J. Physiol. Lung Cell. Mol. Physiol. 291:L426–L435 - PubMed
-
- Becker SD, Bennett M, Stewart JP, Hurst JL. 2007. Serological survey of virus infection among wild house mice (Mus domesticus) in the UK. Lab. Anim. 41:229–238 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
