

The 3' untranslated regions of influenza genomic sequences are 5'PPP-independent ligands for RIG-I
- PMID: 22438882
- PMCID: PMC3305289
- DOI: 10.1371/journal.pone.0032661
The 3' untranslated regions of influenza genomic sequences are 5'PPP-independent ligands for RIG-I
Abstract
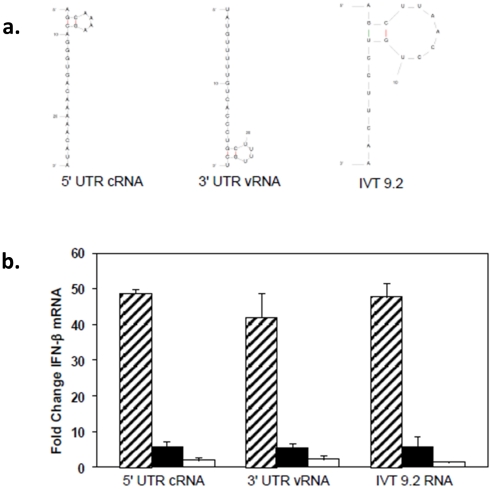
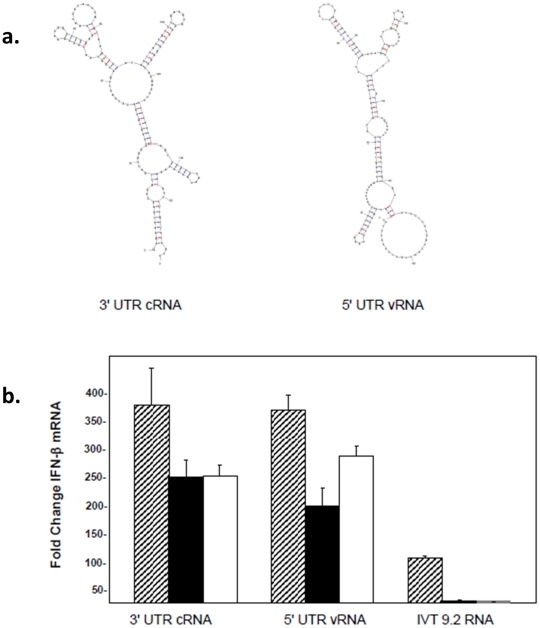
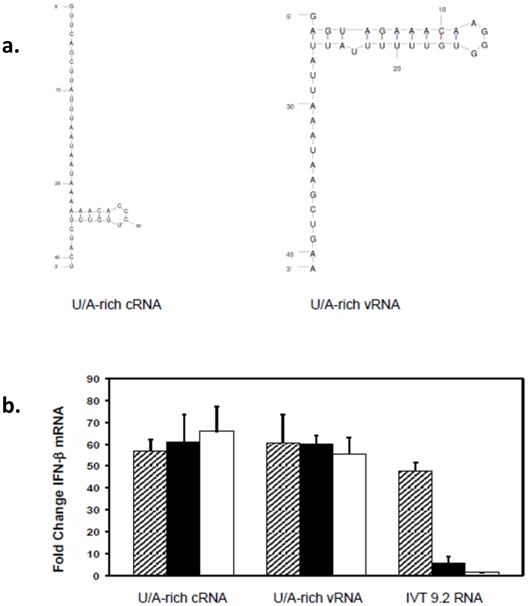
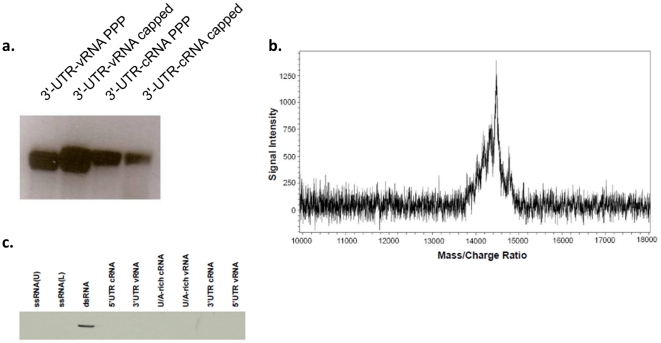
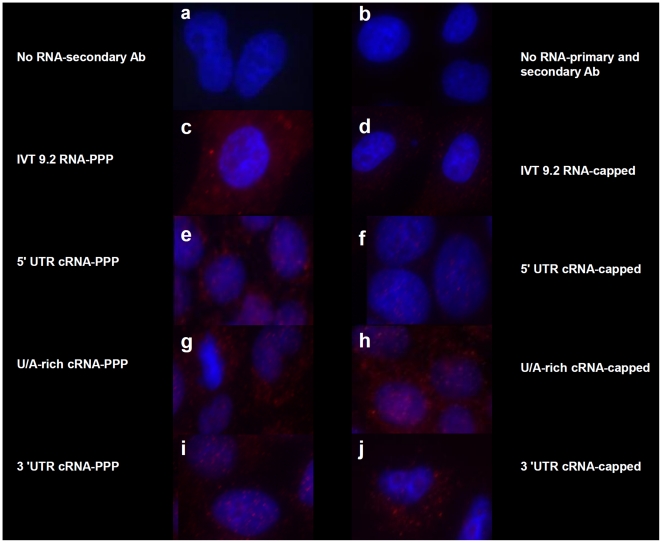
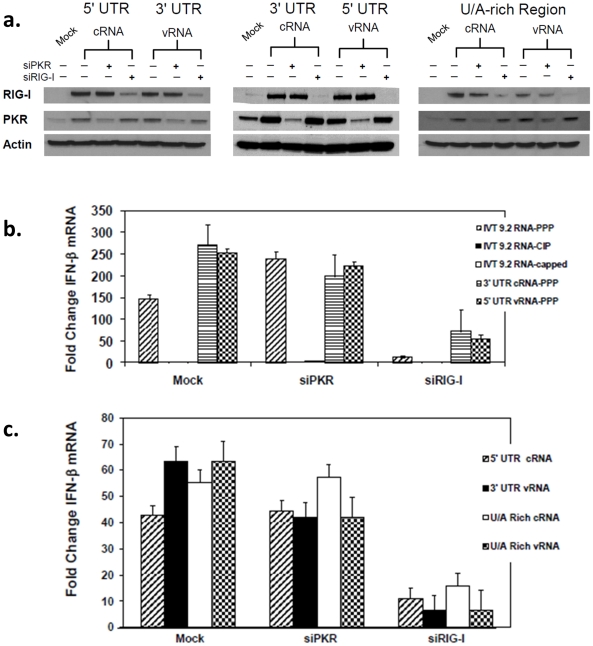
Retinoic acid inducible gene-I (RIG-I) is a key regulator of antiviral immunity. RIG-I is generally thought to be activated by ssRNA species containing a 5'-triphosphate (PPP) group or by unphosphorylated dsRNA up to ~300 bp in length. However, it is not yet clear how changes in the length, nucleotide sequence, secondary structure, and 5' end modification affect the abilities of these ligands to bind and activate RIG-I. To further investigate these parameters in the context of naturally occurring ligands, we examined RNA sequences derived from the 5' and 3' untranslated regions (UTR) of the influenza virus NS1 gene segment. As expected, RIG-I-dependent interferon-β (IFN-β) induction by sequences from the 5' UTR of the influenza cRNA or its complement (26 nt in length) required the presence of a 5'PPP group. In contrast, activation of RIG-I by the 3' UTR cRNA sequence or its complement (172 nt) exhibited only a partial 5'PPP-dependence, as capping the 5' end or treatment with CIP showed a modest reduction in RIG-I activation. Furthermore, induction of IFN-β by a smaller, U/A-rich region within the 3' UTR was completely 5'PPP-independent. Our findings demonstrated that RNA sequence, length, and secondary structure all contributed to whether or not the 5'PPP moiety is needed for interferon induction by RIG-I.
Conflict of interest statement
Figures
References
-
- Akira S. Innate immunity to pathogens: diversity in receptors for microbial recognition. Immunol Rev. 2009;227:5–8. - PubMed
-
- Seth RB, Sun L, Chen ZJ. Antiviral innate immunity pathways. Cell Res. 2006;16:141–147. - PubMed
-
- Diebold SS, Kaisho T, Hemmi H, Akira S, Reis e Sousa C. Innate antiviral responses by means of TLR7-mediated recognition of single-stranded RNA. Science. 2004;303:1529–1531. - PubMed
-
- Alexopoulou L, Holt AC, Medzhitov R, Flavell RA. Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature. 2001;413:732–738. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
