

doi: 10.1186/2045-3701-2-11.
Retinoic acid synthesis and functions in early embryonic development
Affiliations
- PMID: 22439772
- PMCID: PMC3325842
- DOI: 10.1186/2045-3701-2-11
Item in Clipboard
Retinoic acid synthesis and functions in early embryonic development
Cell Biosci.
.
Abstract
Retinoic acid (RA) is a morphogen derived from retinol (vitamin A) that plays important roles in cell growth, differentiation, and organogenesis. The production of RA from retinol requires two consecutive enzymatic reactions catalyzed by different sets of dehydrogenases. The retinol is first oxidized into retinal, which is then oxidized into RA. The RA interacts with retinoic acid receptor (RAR) and retinoic acid X receptor (RXR) which then regulate the target gene expression. In this review, we have discussed the metabolism of RA and the important components of RA signaling pathway, and highlighted current understanding of the functions of RA during early embryonic development.
Figures
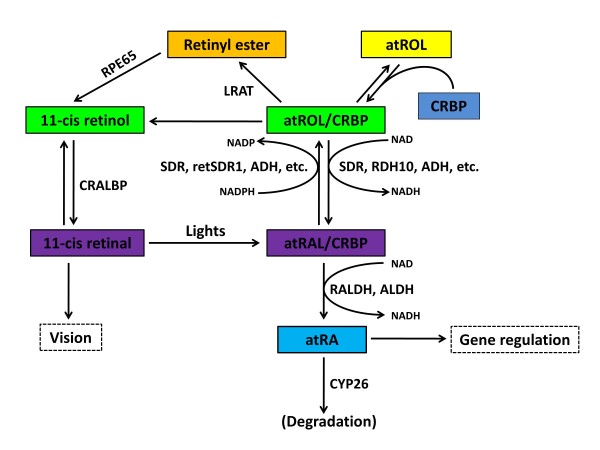
Schematic diagram showing the metabolic pathways of vitamin A metabolism. This illustration lists the major steps of RA metabolism in cell, please refer to the text for details. Abbreviation: NAD, nicotinamide adenine dinucleotide; NADH, The reduced form of NAD; NADP, nicotinamide adenine dinucleotide phosphate; NADPH, The reduced form of NADP; CRBP: cellular retinol-binding protein; LRAT: lecithin retinol acyltransferase; REH: retinyl ester hydrolase; ADH: alcohol dehydrogenase; RDH: retinol dehydrogenase; SDR: short-chain dehydrogenase/reductases; ALDH: aldehyde dehydrogenase; RALDH: retinaldehyde dehydrogenase. CRAD: cis-retinoid/androgen dehydrogenase. Modified from [18,25].
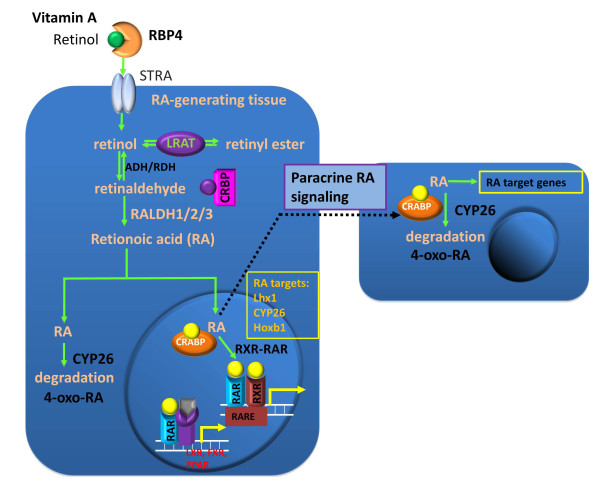
Ilustration of RA and paracrine RA signaling. In serum, retinol is bound to retinol-binding protein 4 (RBP-4) synthesized in the liver. Although retinol is lipid soluble, it enters cells mainly through the interaction with its receptor STRA. In the cell, retinol can either be converted into retinyl esters for storage via lecithin retinol acyltransferase (LRAT) or bind to the cellular retinol binding protein (CRBP). The CRBP-bound retinol is oxidized to retinal by either alcohol dehydrogenase (ADH) or retinol dehydrogenase (RDH), and retinal is oxidized to retinoic acid (RA) by retinaldehyde dehydrogenases (RALDH1/2/3). All-trans retinoic acid (atRA) is the major bioactive component among the retinoids. CYP26 can further oxidize atRA to 4-oxo-RA for degradation. Cellular retinoic acid-binding protein (CRABP) facilitates transportation of atRA into the nucleus where atRA binds its receptors. The ternary complex of ligand-bound RAR and RXR binds to the retinoic acid response element (RARE) and activates the RA target genes. atRA can diffuse to adjacent cells to activate target gene expression in these cells. RAR can also bind to the liver X receptor (LXR), farnesoid X receptor (FXR), and peroxisome proliferator-activated receptor (PPAR) for multiple functions.
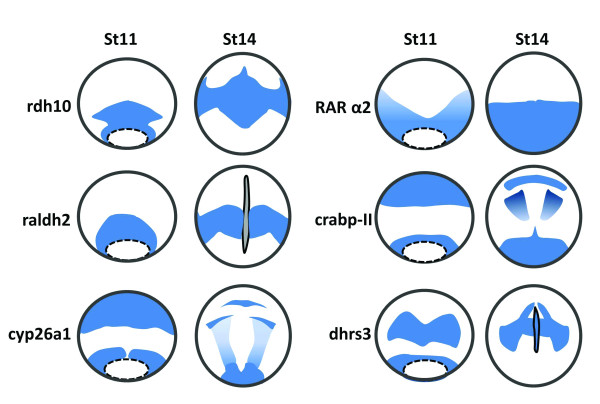
Schematic diagram illustrating expression of the genes that are involved in RA biosynthesis and transportation at gastrula (stage 11) and neurula (stage 14) stages of Xenopus embryos. rdh10 is expressed in the circumblastoporal region of Xenopus gastrula, and the signals at the dorsal side form a zone extending anteriorly. At stage14, the signals are found in trunk paraxial mesoderm region [46]. rdh10 is not expressed in the notochord. raldh2 is expressed in a form of a ring around the vegetal pole with signals more intense in the dorsal blastpore lip. During neurula stages, raldh2 signals are mainly distributed in the trunk paraxial mesoderm, expanding ventrally [38]. rdh10 and raldh2 display overlapping expression pattern in the trunk paraxial mesoderm, with rdh10 expressed more anteriorly than raldh2. cyp26a is expressed in two primary domains at stage11, the posterior domain surrounding the blastopore and the anterior domain covering the anterior part of prospective neural plate. At stage 14, the anterior cyp26a transcripts is developed into three elements corresponding to the cement gland anlage, the mid-/hindbrain boundary, and the auditory placodes, while the posterior expression domain remains in the circumblastoporal area, and the developing neural plate is also covered by a gradient of cyp26a signals with the highest present in posterior region [92]. The expression domains of radlh2 and cyp26a do not overlap at gastrula and neurula stages of Xenopus embryos. rarα2 is expressed in the involuting surface layer surrounding the blastopore, and becomes stronger as the gastrulation proceeds. At stage 14, rarα2 expression is expressed predominantly in the posterior neural plate of the embryos [167]. The expression of the carbp-II is defined into an anterior and a posterior domain at gastrula stages. In the anterior domain, Xenopus crabp-II is limited to the dorsal area which generates prospective head structures. At the neurula stages, crabp-II is expressed in the prospective telencephalon and rhombencephalon, and the most posterior region of the embryos [168]. The dhrs3 signals form a circumblastoporal ring which is similar to rdh10 at stage 11. The signals in the neural plate form two signaling zones and gradually converge towards the midline, forming two signal strips extending posteriorly. In addition, dhrs3 is expressed in the notochord at neurula stages [169]. All these drawings are shown in dorsal view and the blue color represents expression signals.
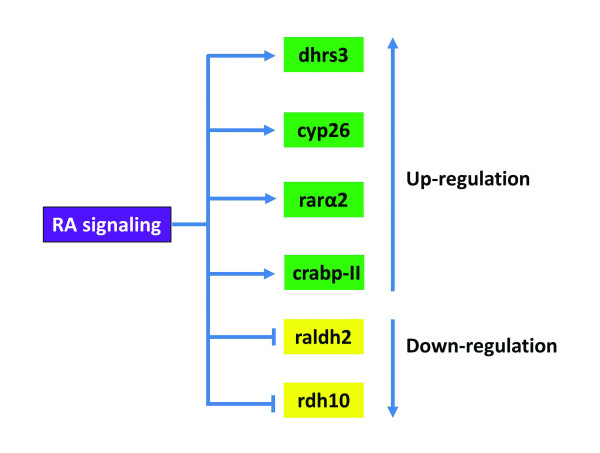
The RA signaling regulates expression of the components involved in RA synthesis and transportation.
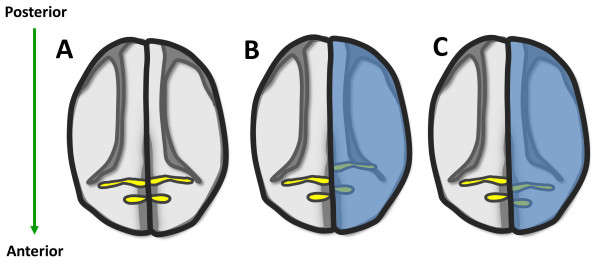
Schematic diagram of krox20 expression in control embryos (A), and in embryos with alterated RA signaling (B, C). Alternation of RA gradient induces shift of the expression domain of krox20. The expression of krox20 labels rhombomeres 3 and 5 (A, yellow). Co-injection of lacZ and cyp26 or rdh10 or rdh2 mRNA into one blastomere of two-cell stage Xenopus embryos is indicated by LacZ staining (B, C, blue). The uninjected side is served as control. Ectopic cyp26 at one side of the embryos causes posterior shift of krox20 (B, yellow). In contrast to cyp26, overexpression of rdh10 and radlh2 leads to anterior shift of krox20 (C, yellow). This diagram is based on the previous reports and our unpublished data [46,92,129] (RKT Kam, Y Chen, WY Chan and H Zhao. Dhrs3 attenuates the retinoic acid signaling and is required for early embryonic patterning. Submitted.).
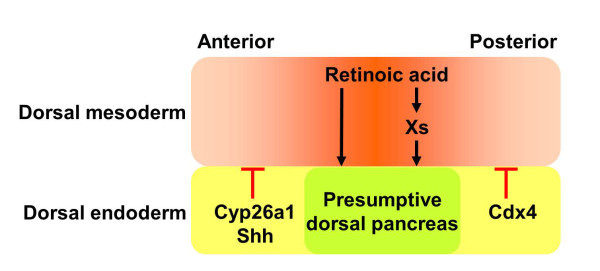
RA plays an essential role in dorsal pancreas specification. During gastrulation and early neurulation, the field in dorsal endoderm that gives rise to the presumptive dorsal pancreas is just underlying the dorsal mesoderm where the highest concentration of RA is generated. These RA signals can act to underly endoderm either directly, or indirectly via first activating unknown factors (Xs) in mesoderm, which then act on the endoderm for dorsal pancreas formation. As described in the text, Shh and Cyp26a1 expressed in more anterior dorsal endoderm block pancreas formation probably via inhibiting the RA signaling activity or directly degrading RA. Cdx4 expressed in the posterior dorsal endoderm helps to establish the posterior boundary of dorsal pancreas via antagonizing the RA signaling.
References
-
- Duriancik DM, Lackey DE, Hoag KA. Vitamin A as a regulator of antigen presenting cells. J Nutr. pp. 1395–1399. - PubMed
LinkOut - more resources
Full Text Sources
