

Macroevolution of venom apparatus innovations in auger snails (Gastropoda; Conoidea; Terebridae)
- PMID: 22440724
- PMCID: PMC3389042
- DOI: 10.1016/j.ympev.2012.03.001
Macroevolution of venom apparatus innovations in auger snails (Gastropoda; Conoidea; Terebridae)
Abstract
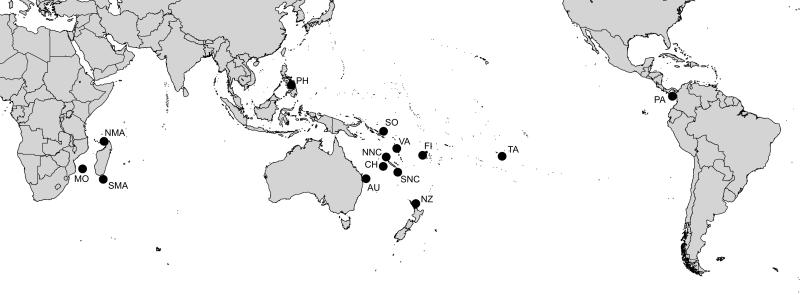
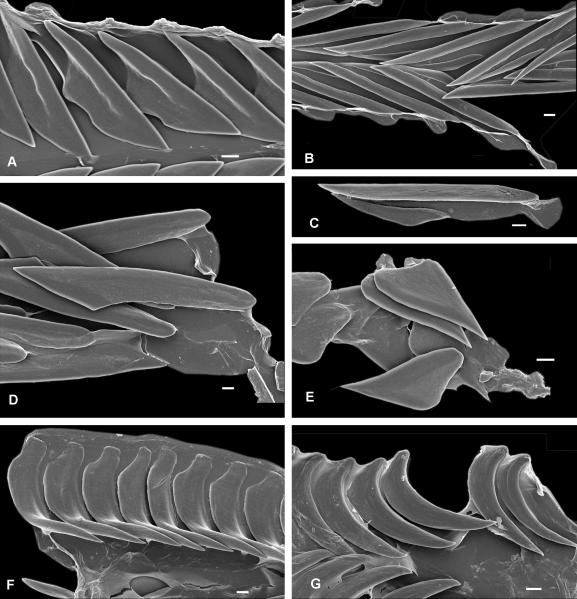
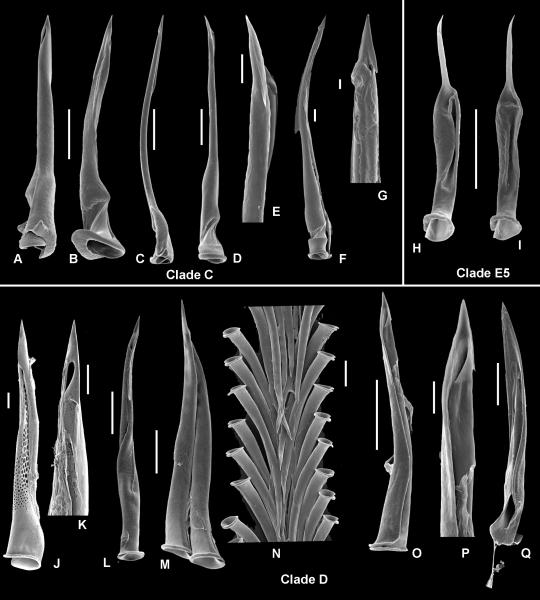
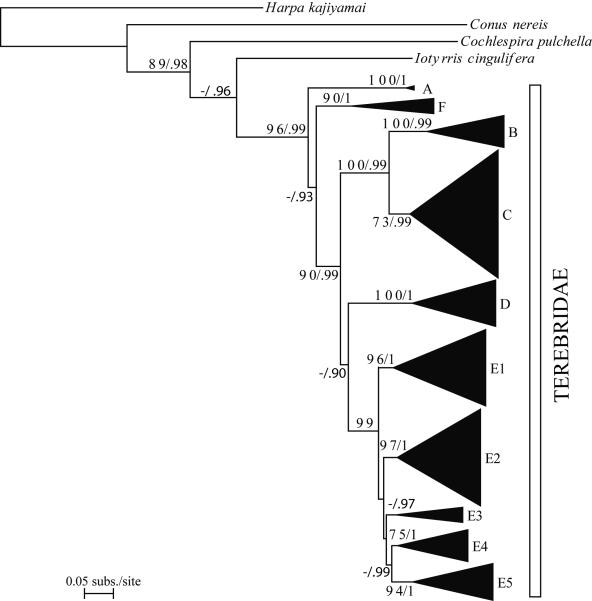
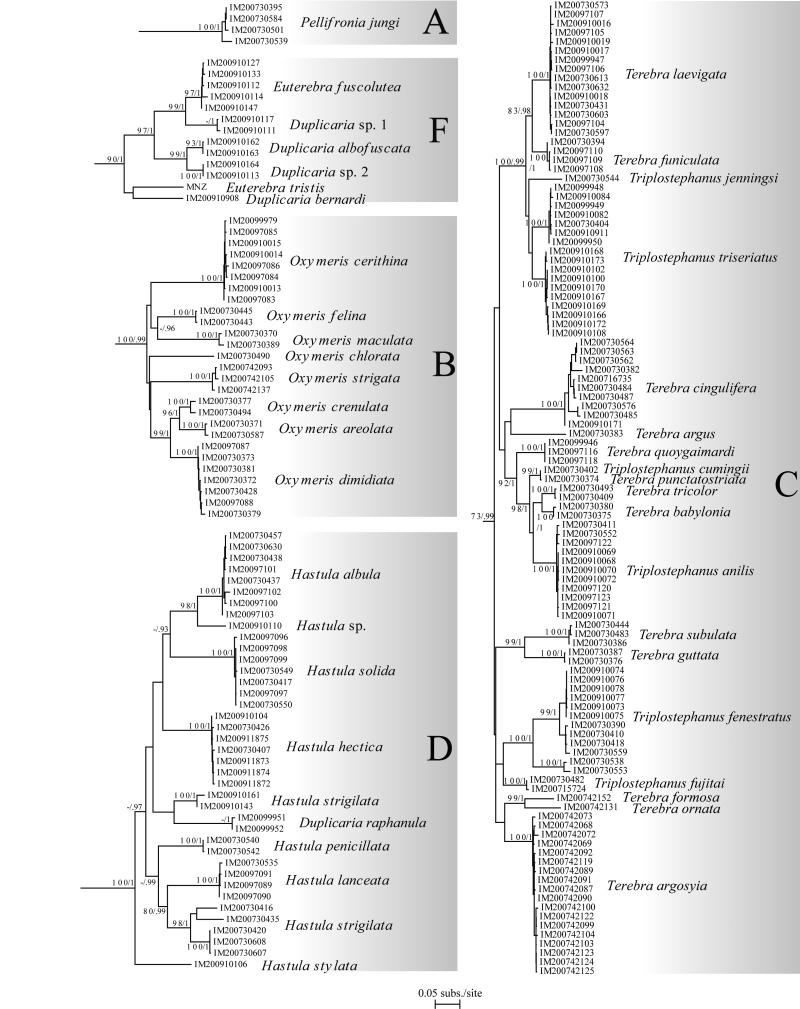
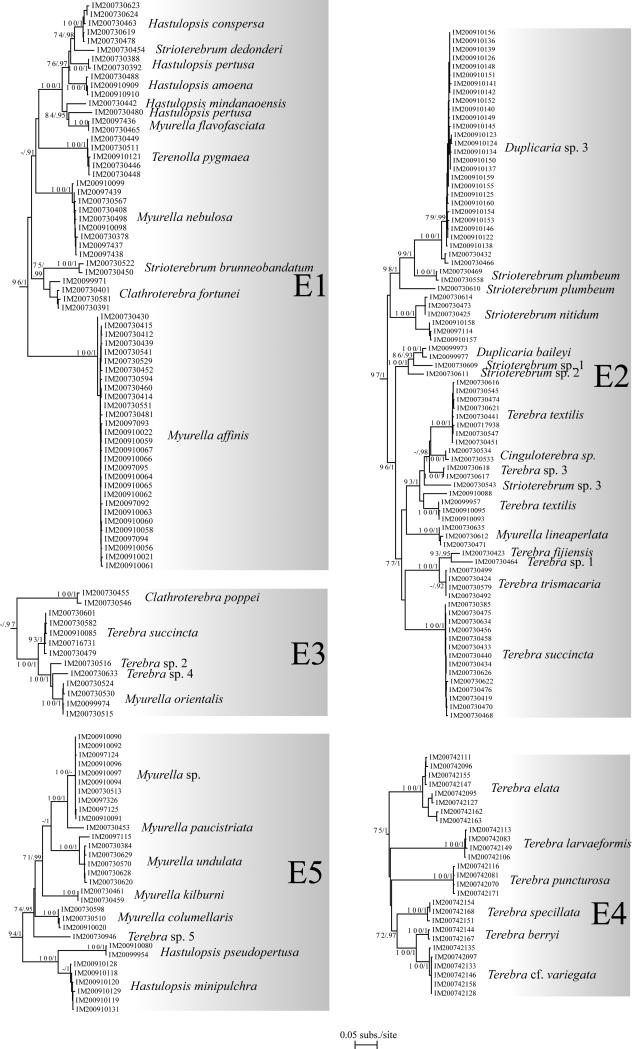
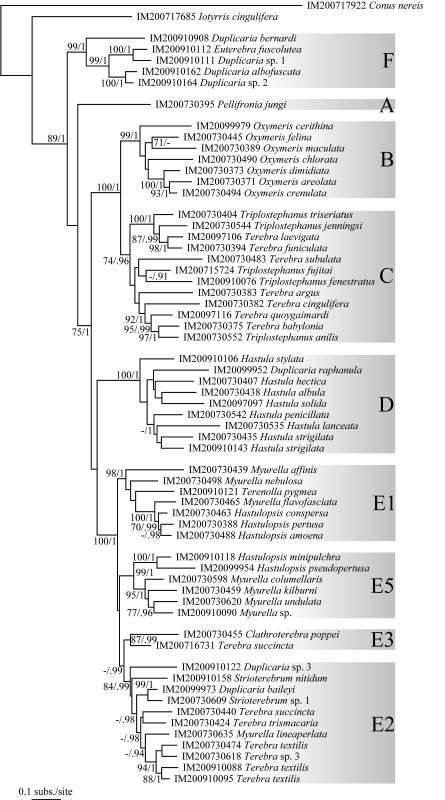
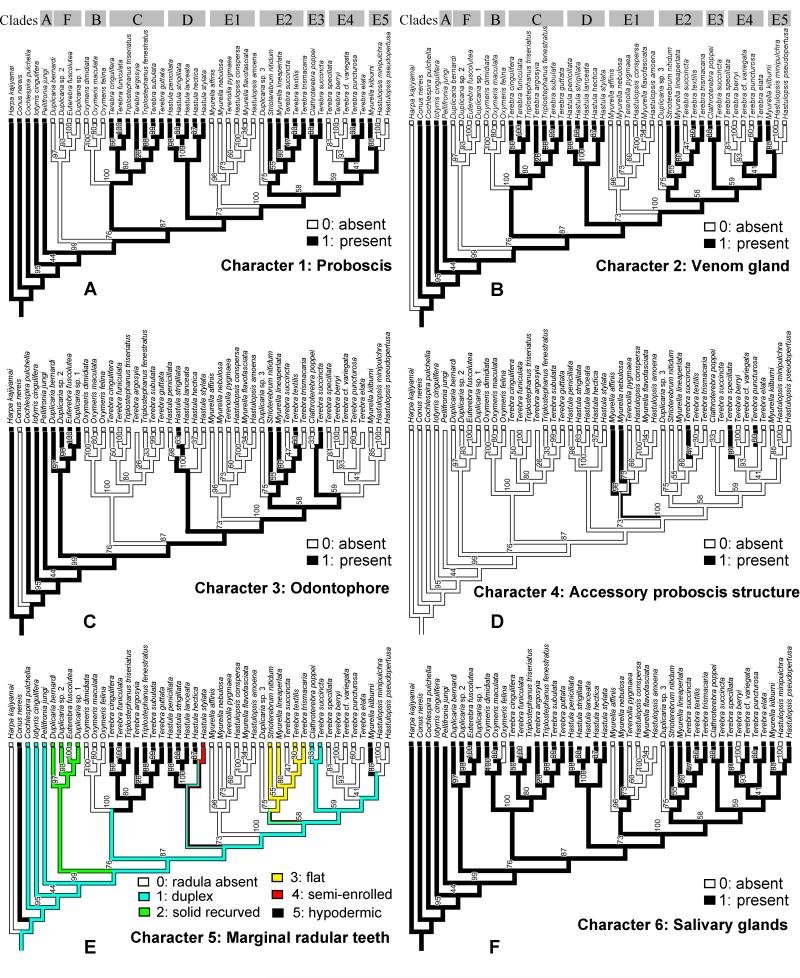
The Terebridae are a diverse family of tropical and subtropical marine gastropods that use a complex and modular venom apparatus to produce toxins that capture polychaete and enteropneust preys. The complexity of the terebrid venom apparatus suggests that venom apparatus development in the Terebridae could be linked to the diversification of the group and can be analyzed within a molecular phylogenetic scaffold to better understand terebrid evolution. Presented here is a molecular phylogeny of 89 terebrid species belonging to 12 of the 15 currently accepted genera, based on Bayesian inference and Maximum Likelihood analyses of amplicons of 3 mitochondrial (COI, 16S and 12S) and one nuclear (28S) genes. The evolution of the anatomy of the terebrid venom apparatus was assessed by mapping traits of six related characters: proboscis, venom gland, odontophore, accessory proboscis structure, radula, and salivary glands. A novel result concerning terebrid phylogeny was the discovery of a previously unrecognized lineage, which includes species of Euterebra and Duplicaria. The non-monophyly of most terebrid genera analyzed indicates that the current genus-level classification of the group is plagued with homoplasy and requires further taxonomic investigations. Foregut anatomy in the family Terebridae reveals an inordinate diversity of features that covers the range of variability within the entire superfamily Conoidea, and that hypodermic radulae have likely evolved independently on at least three occasions. These findings illustrate that terebrid venom apparatus evolution is not perfunctory, and involves independent and numerous changes of central features in the foregut anatomy. The multiple emergence of hypodermic marginal radular teeth in terebrids are presumably associated with variable functionalities, suggesting that terebrids have adapted to dietary changes that may have resulted from predator-prey relationships. The anatomical and phylogenetic results presented serve as a starting point to advance investigations about the role of predator-prey interactions in the diversification of the Terebridae and the impact on their peptide toxins, which are promising bioactive compounds for biomedical research and therapeutic drug development.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures

References
-
- Barnosky AD. Distinguishing the effects of the Red Queen and Court Jester on Miocene mammal evolution in the northern Rocky Mountains. J. Vertebr. Paleontol. 2001;21:172–185.
-
- Berenbaum M. Coumarins and caterpillars: a case for coevolution. Evolution. 1983;37:163–179. - PubMed
-
- Berenbaum M, Feeny P. Toxicity of angular furanocoumarins to swallowtail butterflies: escalation in a coevolutionary arms race? Science. 1981;212:927–929. - PubMed
-
- Biardi JE, Chien DC, Coss RG. California ground squirrel (Spermophilus beecheyi) defenses against rattlesnake venom digestive and hemostatic toxins. J. Chem. Ecol. 2005;31:2501–2518. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
