

Targeted deletion of ERK5 MAP kinase in the developing nervous system impairs development of GABAergic interneurons in the main olfactory bulb and behavioral discrimination between structurally similar odorants
- PMID: 22442076
- PMCID: PMC3333842
- DOI: 10.1523/JNEUROSCI.6260-11.2012
Targeted deletion of ERK5 MAP kinase in the developing nervous system impairs development of GABAergic interneurons in the main olfactory bulb and behavioral discrimination between structurally similar odorants
Abstract
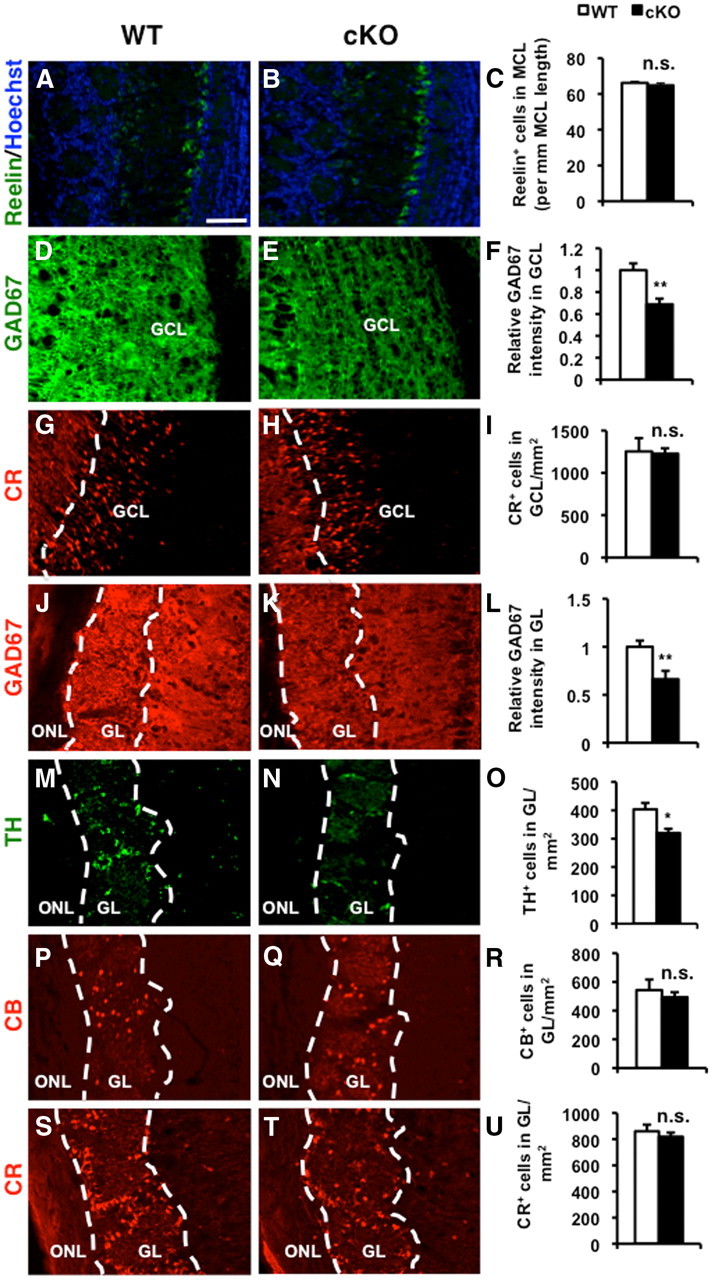
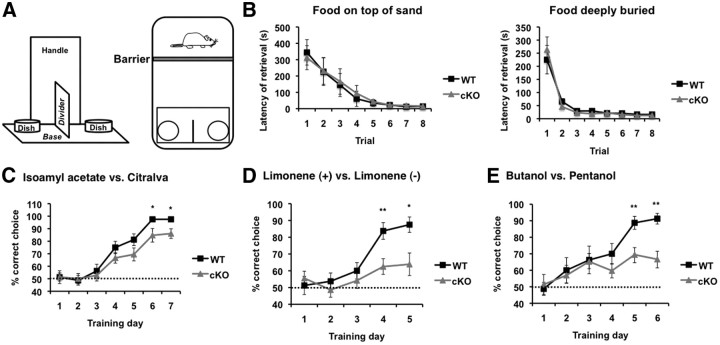
ERK5 MAP kinase is highly expressed in the developing nervous system and has been implicated in promoting the survival of immature neurons in culture. However, its role in the development and function of the mammalian nervous system has not been established in vivo. Here, we report that conditional deletion of the erk5 gene in mouse neural stem cells during development reduces the number of GABAergic interneurons in the main olfactory bulb (OB). Our data suggest that this is due to a decrease in proliferation and an increase in apoptosis in the subventricular zone and rostral migratory stream of ERK5 mutant mice. Interestingly, ERK5 mutant mice have smaller OB and are impaired in odor discrimination between structurally similar odorants. We conclude that ERK5 is a novel signaling pathway regulating developmental OB neurogenesis and olfactory behavior.
Figures









References
-
- Adam Y, Mizrahi A. Circuit formation and maintenance–perspectives from the mammalian olfactory bulb. Curr Opin Neurobiol. 2010;20:134–140. - PubMed
-
- Allen ZJ, 2nd, Waclaw RR, Colbert MC, Campbell K. Molecular identity of olfactory bulb interneurons: transcriptional codes of periglomerular neuron subtypes. J Mol Histol. 2007;38:517–525. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous