

P2X2 and P2X5 subunits define a new heteromeric receptor with P2X7-like properties
- PMID: 22442090
- PMCID: PMC6621234
- DOI: 10.1523/JNEUROSCI.6332-11.2012
P2X2 and P2X5 subunits define a new heteromeric receptor with P2X7-like properties
Abstract
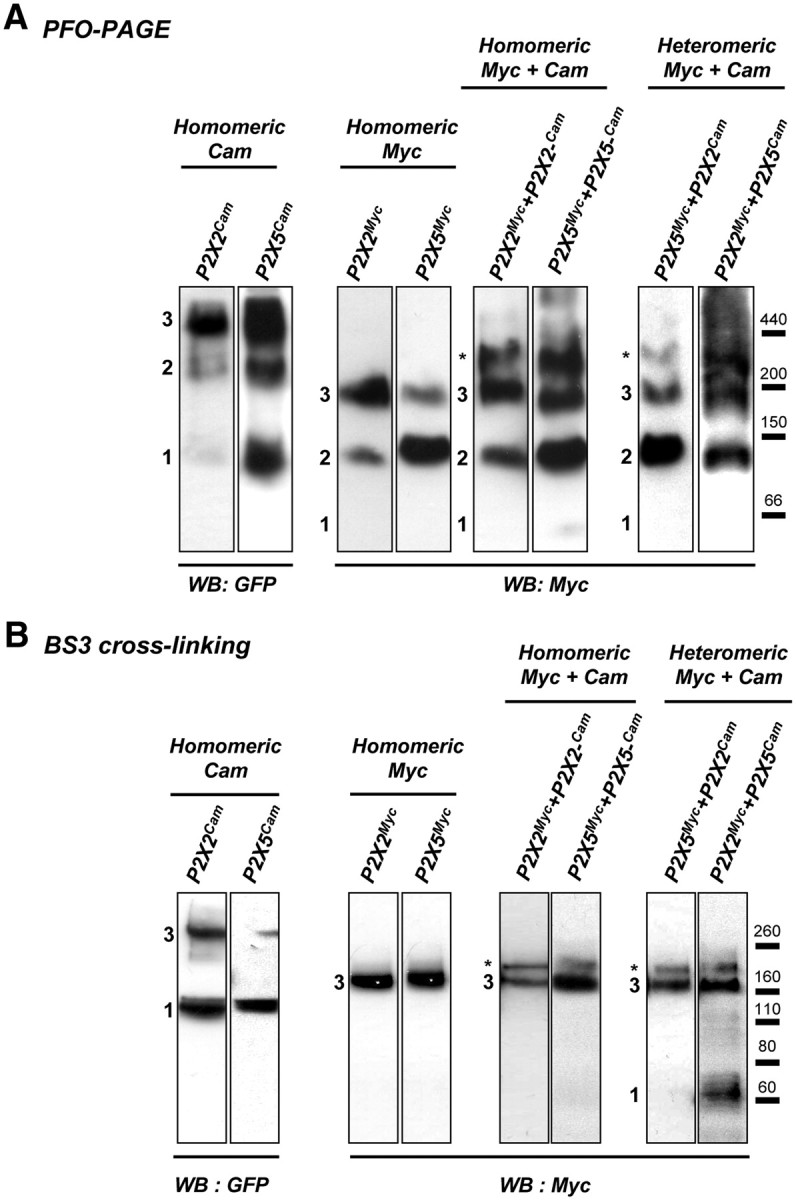
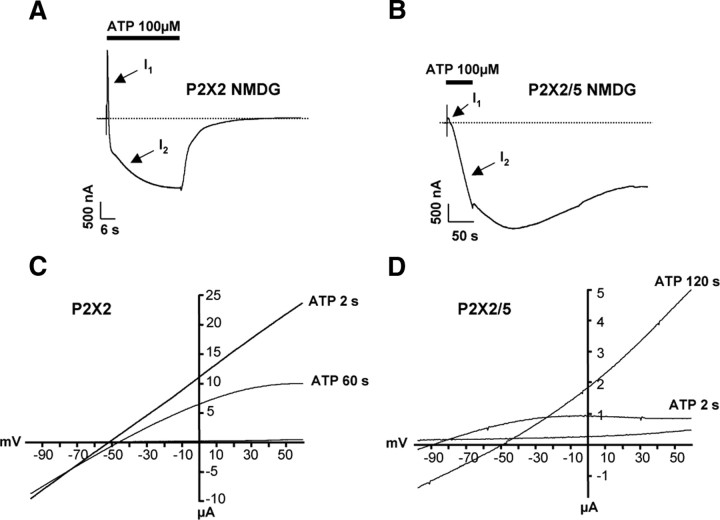
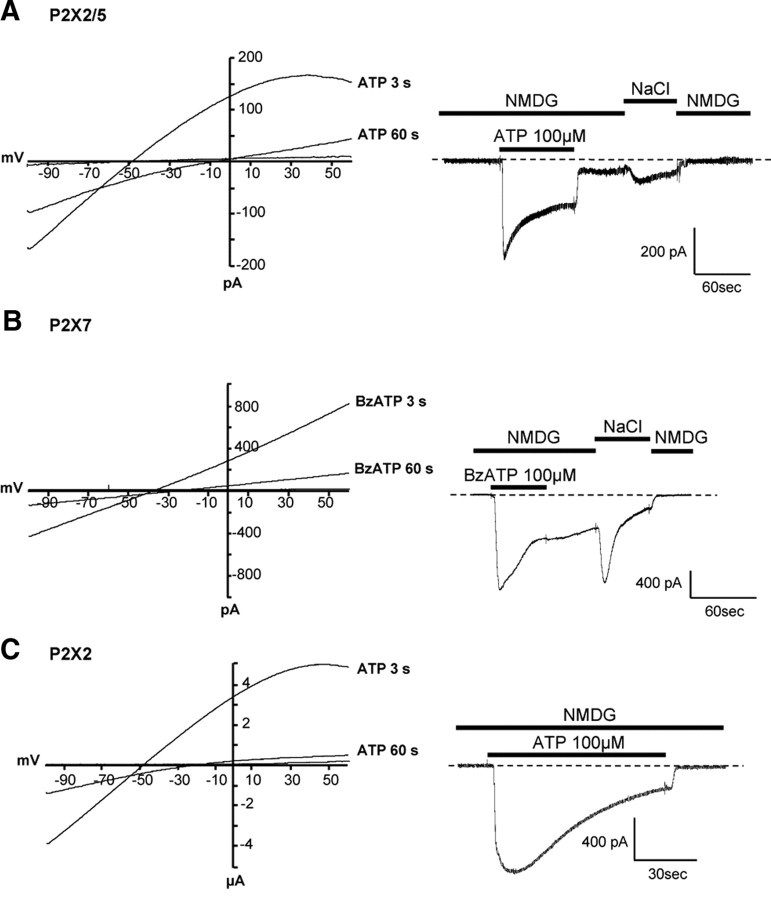
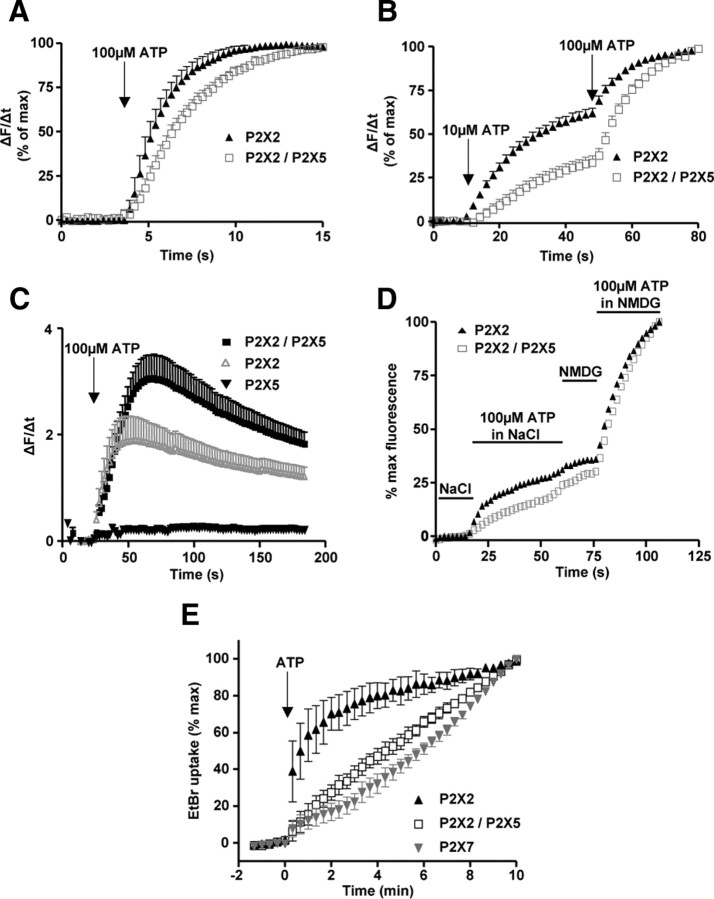
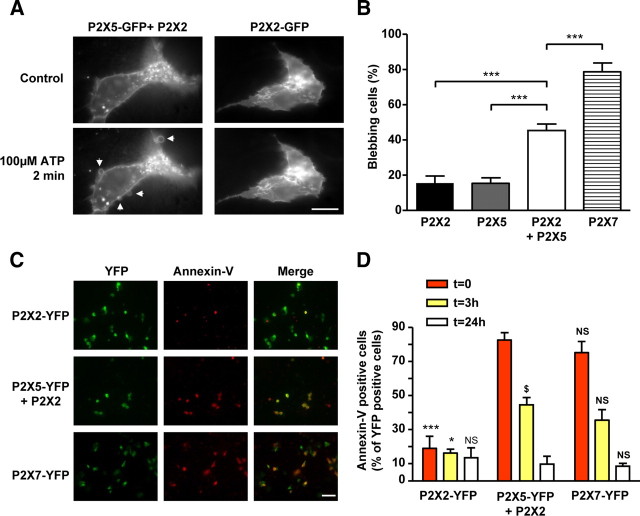
Ligand-gated ion channels are prototypic oligomeric membrane proteins whose stoichiometry determines their functional properties and subcellular localization. Deciphering the quaternary structure of such protein complexes is an arduous task and usually requires the combination of multiple approaches. ATP-gated P2X receptors are formed by the association of three subunits, but the quaternary arrangement of the seven P2X subunits at the plasma membrane remains poorly characterized. By combining bioluminescence resonance energy transfer, bifunctional fluorescence complementation and protein biochemistry, we developed an experimental approach that allows precise determination of rat P2X receptor quaternary assembly. We found that P2X5 subunits associate with P2X1, P2X2, and P2X4 subunits. We demonstrate that P2X5 and P2X2 subunits interact to form as yet uncharacterized heteromeric receptors with alternate stoichiometries, both present at the plasma membrane. P2X2/5 receptors display functional properties such as pore dilatation, membrane blebbing, and phosphatidylserine exposure that were previously thought to be characteristic hallmarks of the P2X7 receptor. In mouse, P2X2 and P2X5 subunits colocalize and physically interact in specific neuronal populations suggesting that other P2X receptors might contribute to cellular responses typically attributed to P2X7 receptor.
Figures
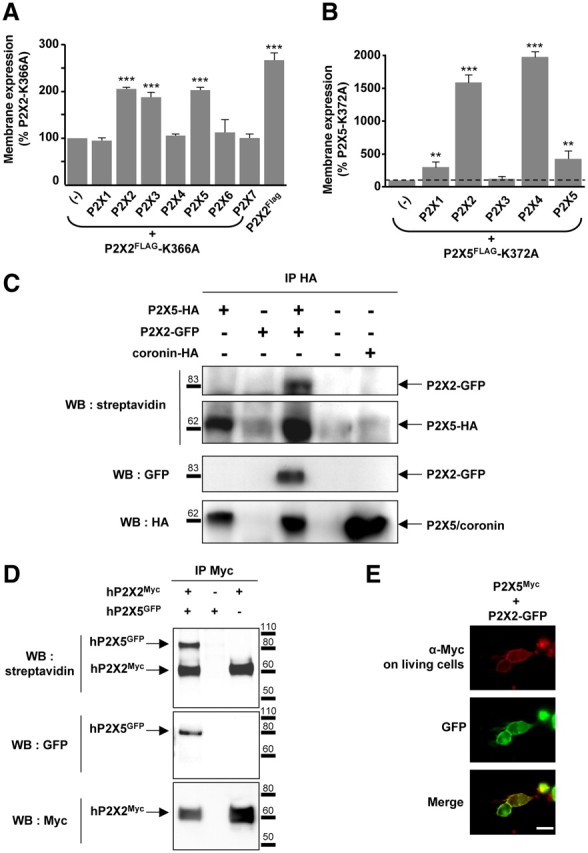
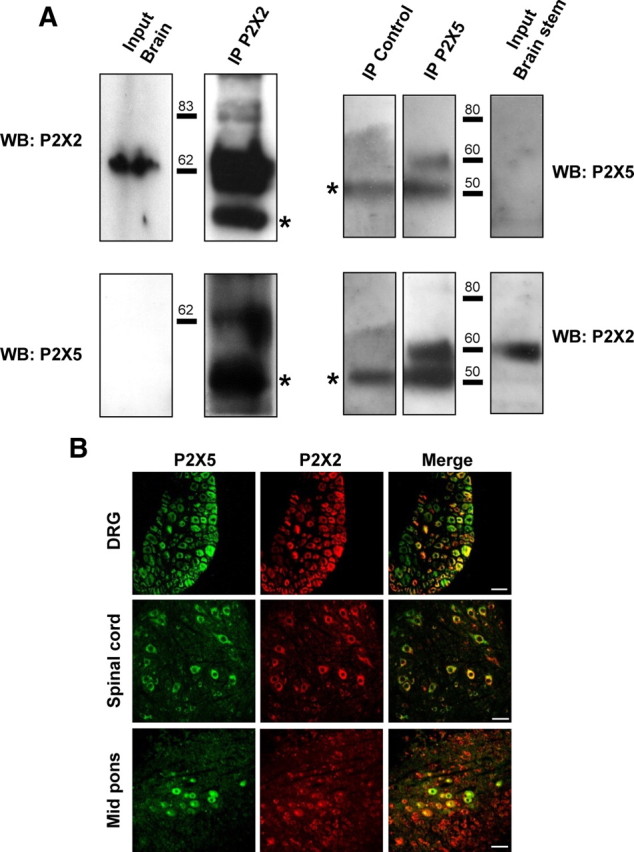
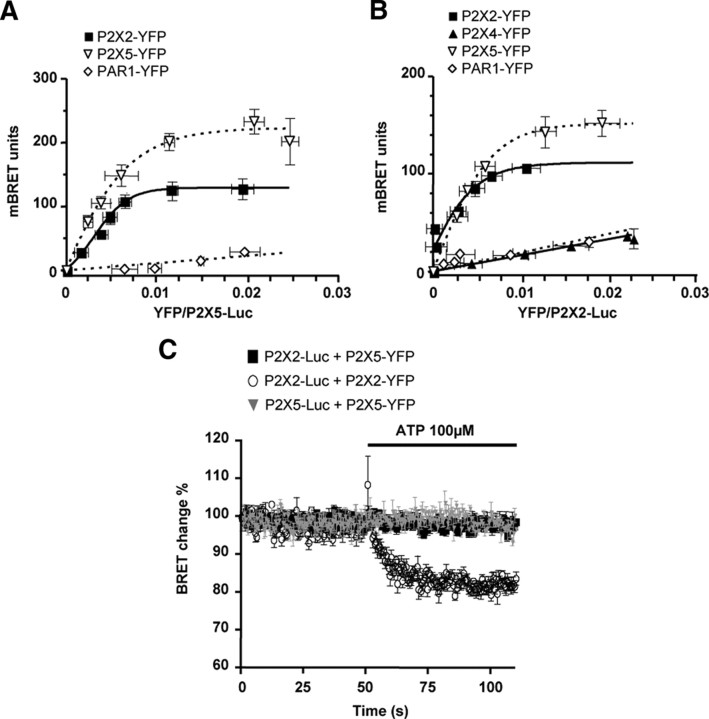
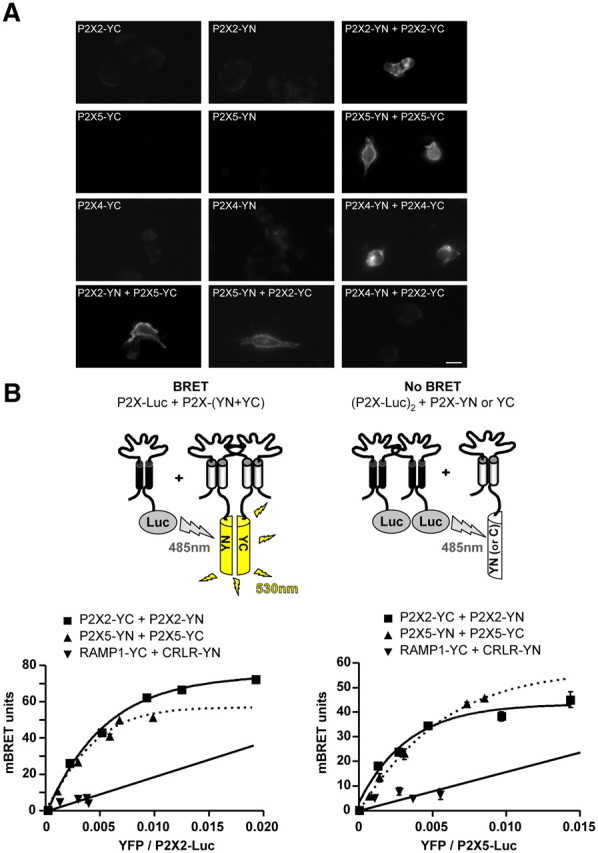
References
-
- Anderson CM, Nedergaard M. Emerging challenges of assigning P2X7 receptor function and immunoreactivity in neurons. Trends Neurosci. 2006;29:257–262. - PubMed
-
- Aschrafi A, Sadtler S, Niculescu C, Rettinger J, Schmalzing G. Trimeric achitecture of homomeric P2X2 and heteromeric P2X1+2 receptor subtypes. J Mol Biol. 2004;342:333–343. - PubMed
-
- Ayoub MA, Maurel D, Binet V, Fink M, Prézeau L, Ansanay H, Pin JP. Real-time analysis of agonist-induced activation of protease-activated receptor 1/Galphai1 protein complex measured by bioluminescence resonance energy transfer in living cells. Mol Pharmacol. 2007;71:1329–1340. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases