

Role of mitochondrial Ca2+ in the regulation of cellular energetics
- PMID: 22443365
- PMCID: PMC3332087
- DOI: 10.1021/bi2018909
Role of mitochondrial Ca2+ in the regulation of cellular energetics
Abstract
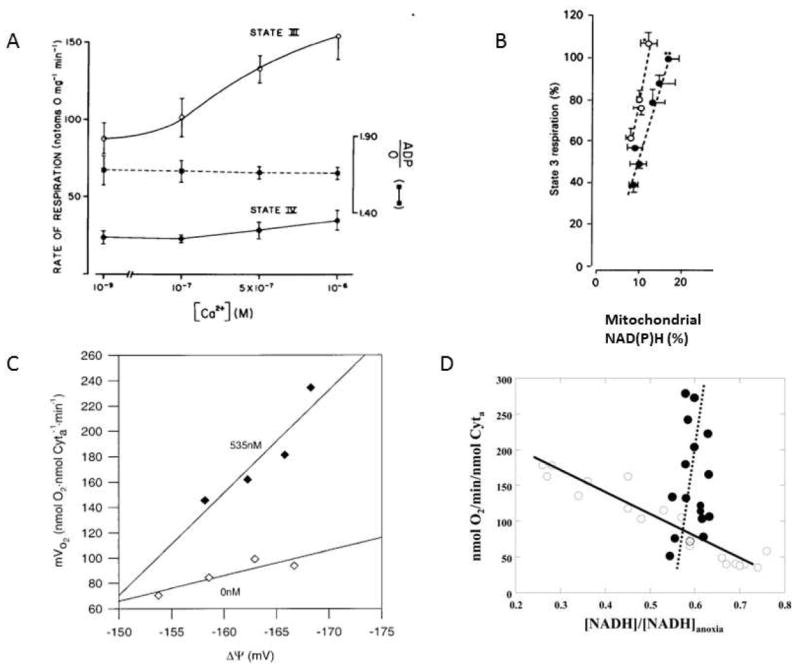
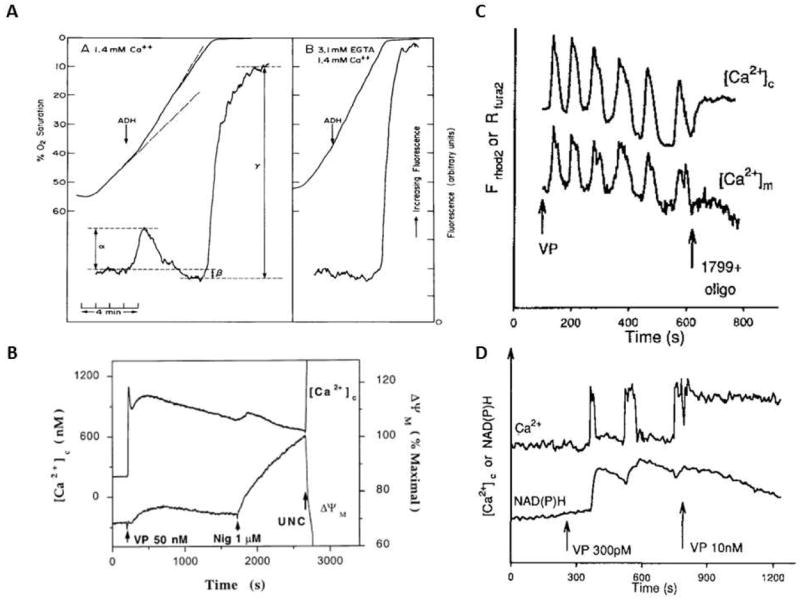
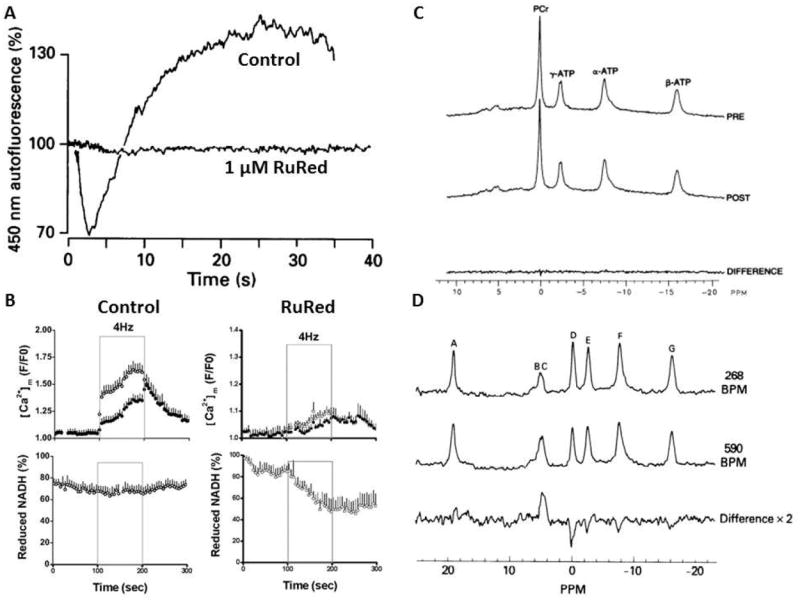
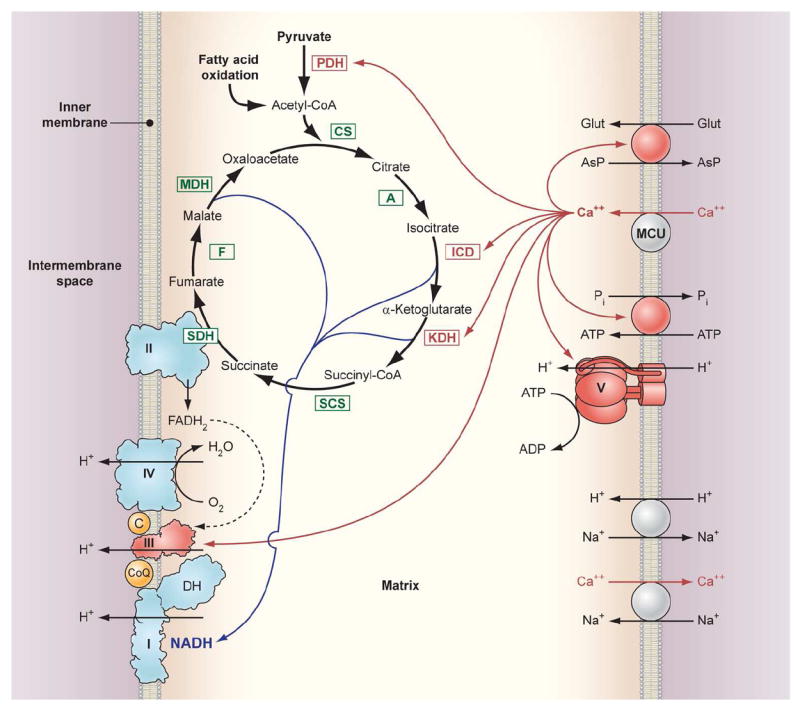
Calcium is an important signaling molecule involved in the regulation of many cellular functions. The large free energy in the Ca(2+) ion membrane gradients makes Ca(2+) signaling inherently sensitive to the available cellular free energy, primarily in the form of ATP. In addition, Ca(2+) regulates many cellular ATP-consuming reactions such as muscle contraction, exocytosis, biosynthesis, and neuronal signaling. Thus, Ca(2+) becomes a logical candidate as a signaling molecule for modulating ATP hydrolysis and synthesis during changes in numerous forms of cellular work. Mitochondria are the primary source of aerobic energy production in mammalian cells and also maintain a large Ca(2+) gradient across their inner membrane, providing a signaling potential for this molecule. The demonstrated link between cytosolic and mitochondrial Ca(2+) concentrations, identification of transport mechanisms, and the proximity of mitochondria to Ca(2+) release sites further supports the notion that Ca(2+) can be an important signaling molecule in the energy metabolism interplay of the cytosol with the mitochondria. Here we review sites within the mitochondria where Ca(2+) plays a role in the regulation of ATP generation and potentially contributes to the orchestration of cellular metabolic homeostasis. Early work on isolated enzymes pointed to several matrix dehydrogenases that are stimulated by Ca(2+), which were confirmed in the intact mitochondrion as well as cellular and in vivo systems. However, studies in these intact systems suggested a more expansive influence of Ca(2+) on mitochondrial energy conversion. Numerous noninvasive approaches monitoring NADH, mitochondrial membrane potential, oxygen consumption, and workloads suggest significant effects of Ca(2+) on other elements of NADH generation as well as downstream elements of oxidative phosphorylation, including the F(1)F(O)-ATPase and the cytochrome chain. These other potential elements of Ca(2+) modification of mitochondrial energy conversion will be the focus of this review. Though most specific molecular mechanisms have yet to be elucidated, it is clear that Ca(2+) provides a balanced activation of mitochondrial energy metabolism that exceeds the alteration of dehydrogenases alone.
This article not subject to U.S. Copyright. Published 2012 by the American Chemical Society
Figures
References
-
- Hasselbach W, Oetliker H. Energetics and electrogenicity of the sarcoplasmic reticulum calcium pump. Annu Rev Physiol. 1983;45:325–339. - PubMed
-
- Chen W, London R, Murphy E, Steenbergen C. Regulation of the Ca2+ gradient across the sarcoplasmic reticulum in perfused rabbit heart. A 19F nuclear magnetic resonance study. Circ Res. 1998;83:898–907. - PubMed
-
- Tian R, Halow JM, Meyer M, Dillmann WH, Figueredo VM, Ingwall JS, Camacho SA. Thermodynamic limitation for Ca2+ handling contributes to decreased contractile reserve in rat hearts. Am J Physiol. 1998;275:H2064–H2071. - PubMed
-
- Hill AV. A challange to biochemists. Biochim Biophys Acta. 1950;4:4–11. - PubMed
-
- Devin A, Rigoulet M. Mechanisms of mitochondrial response to variations in energy demand in eukaryotic cells. Am J Physiol Cell Physiol. 2007;292:C52–C58. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
