

Growth cone travel in space and time: the cellular ensemble of cytoskeleton, adhesion, and membrane
- PMID: 22445336
- PMCID: PMC3319373
- DOI: 10.1016/j.neuron.2012.03.005
Growth cone travel in space and time: the cellular ensemble of cytoskeleton, adhesion, and membrane
Abstract
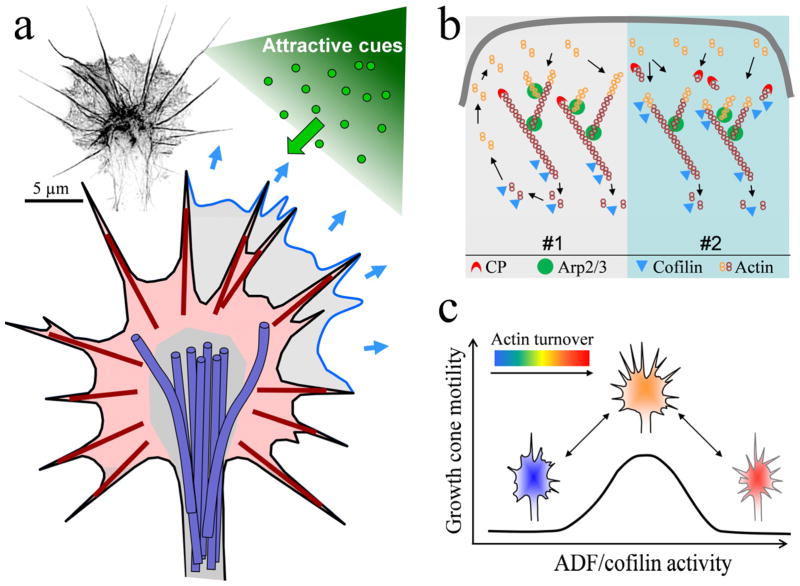
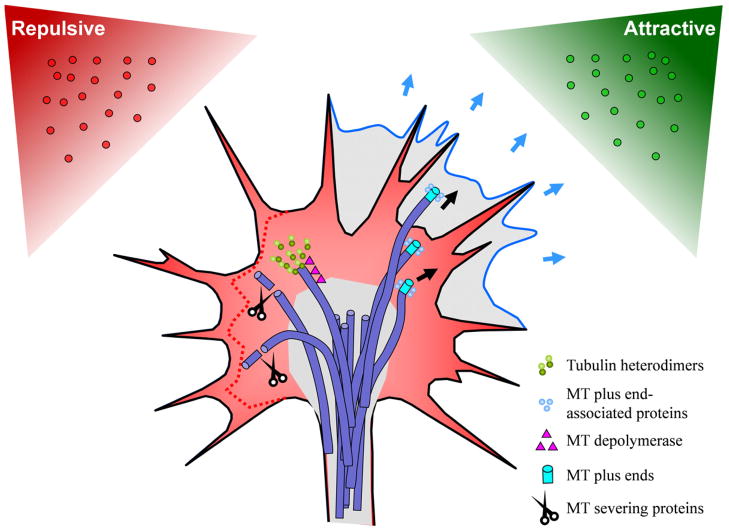
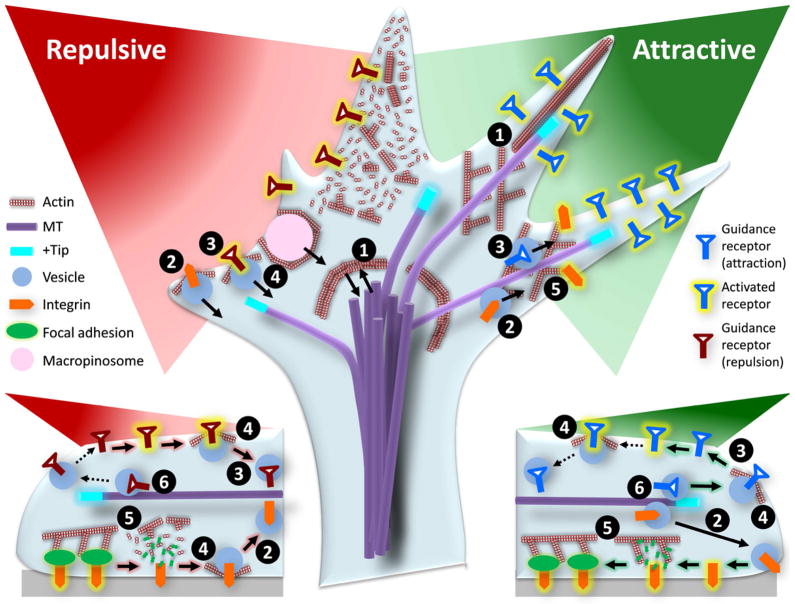
Growth cones, found at the tip of axonal projections, are the sensory and motile organelles of developing neurons that enable axon pathfinding and target recognition for precise wiring of the neural circuitry. To date, many families of conserved guidance molecules and their corresponding receptors have been identified that work in space and time to ensure billions of axons to reach their targets. Research in the past two decades has also gained significant insight into the ways in which growth cones translate extracellular signals into directional migration. This review aims to examine new progress toward understanding the cellular mechanisms underlying directional motility of the growth cone and to discuss questions that remain to be addressed. Specifically, we will focus on the cellular ensemble of cytoskeleton, adhesion, and membrane and examine how the intricate interplay between these processes orchestrates the directed movement of growth cones.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Aizawa H, Wakatsuki S, Ishii A, Moriyama K, Sasaki Y, Ohashi K, Sekine-Aizawa Y, Sehara-Fujisawa A, Mizuno K, Goshima Y, Yahara I. Phosphorylation of cofilin by LIM-kinase is necessary for semaphorin 3A-induced growth cone collapse. Nat Neurosci. 2001;4:367–373. - PubMed
-
- Andrianantoandro E, Pollard TD. Mechanism of actin filament turnover by severing and nucleation at different concentrations of ADF/cofilin. Mol Cell. 2006;24:13–23. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
