

Depleting components of the THO complex causes increased telomere length by reducing the expression of the telomere-associated protein Rif1p
- PMID: 22448247
- PMCID: PMC3308969
- DOI: 10.1371/journal.pone.0033498
Depleting components of the THO complex causes increased telomere length by reducing the expression of the telomere-associated protein Rif1p
Erratum in
- PLoS One. 2012;7(5): doi/10.1371/annotation/2e35aeeb-4f49-4957-931f-9d942a856e42
Abstract
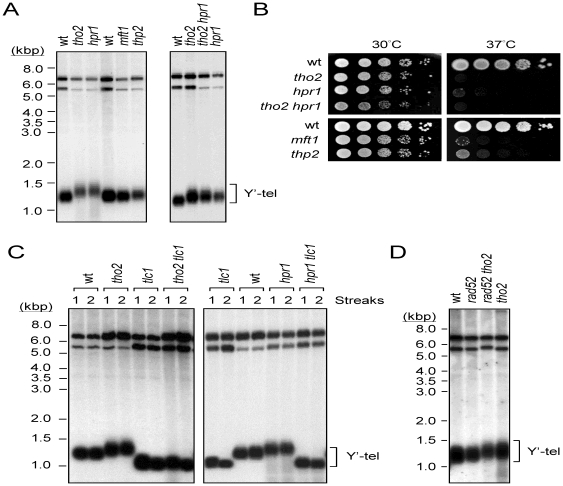
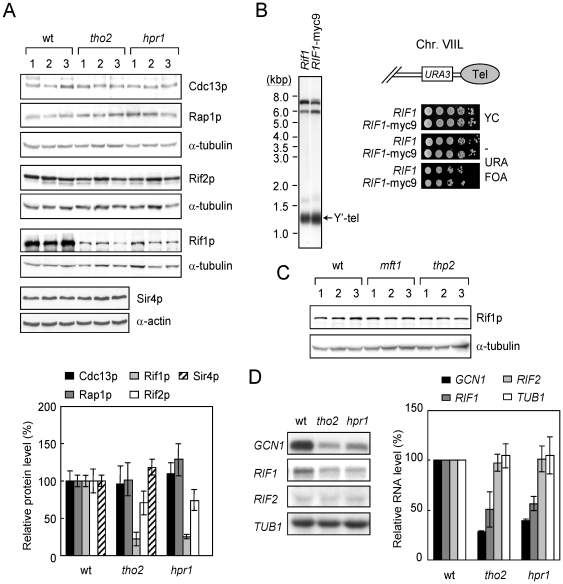
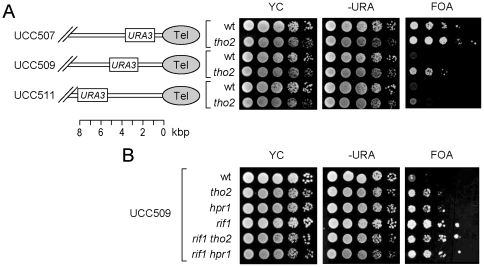
Telomere length is regulated mostly by proteins directly associated with telomeres. However, genome-wide analysis of Saccharomyces cerevisiae mutants has revealed that deletion of Hpr1p, a component of the THO complex, also affects telomere length. The THO complex comprises four protein subunits, namely, Tho2p, Hpr1p, Mft1p, and Thp2p. These subunits interplay between transcription elongation and co-transcriptional assembly of export-competent mRNPs. Here we found that the deletion of tho2 or hpr1 caused telomere lengthening by ∼50-100 bps, whereas that of mft1 or thp2 did not affect telomere length. Since the THO complex functions in transcription elongation, we analyzed the expression of telomere-associated proteins in mutants depleted of complex components. We found that both the mRNA and protein levels of RIF1 were decreased in tho2 and hpr1 cells. RIF1 encodes a 1917-amino acid polypeptide that is involved in regulating telomere length and the formation of telomeric heterochromatin. Hpr1p and Tho2p appeared to affect telomeres through Rif1p, as increased Rif1p levels suppressed the telomere lengthening in tho2 and hpr1 cells. Moreover, yeast cells carrying rif1 tho2 or rif1 hpr1 double mutations showed telomere lengths and telomere silencing effects similar to those observed in the rif1 mutant. Thus, we conclude that mutations of components of the THO complex affect telomere functions by reducing the expression of a telomere-associated protein, Rif1p.
Conflict of interest statement
Figures


Similar articles
-
A novel Rap1p-interacting factor, Rif2p, cooperates with Rif1p to regulate telomere length in Saccharomyces cerevisiae.Genes Dev. 1997 Mar 15;11(6):748-60. doi: 10.1101/gad.11.6.748. Genes Dev. 1997. PMID: 9087429
-
The yeast THO complex and mRNA export factors link RNA metabolism with transcription and genome instability.EMBO J. 2002 Jul 1;21(13):3526-35. doi: 10.1093/emboj/cdf335. EMBO J. 2002. PMID: 12093753 Free PMC article.
-
Architecture and nucleic acids recognition mechanism of the THO complex, an mRNP assembly factor.EMBO J. 2012 Mar 21;31(6):1605-16. doi: 10.1038/emboj.2012.10. Epub 2012 Feb 7. EMBO J. 2012. PMID: 22314234 Free PMC article.
-
Beyond interacting with Rap1: Dissecting the roles of Rif1.Int J Biol Macromol. 2025 May;306(Pt 3):141560. doi: 10.1016/j.ijbiomac.2025.141560. Epub 2025 Mar 1. Int J Biol Macromol. 2025. PMID: 40032092 Review.
-
Rap1p and telomere length regulation in yeast.Ciba Found Symp. 1997;211:76-93; discussion 93-103. doi: 10.1002/9780470515433.ch6. Ciba Found Symp. 1997. PMID: 9524752 Review.
Cited by
-
Telomeric transcripts stimulate telomere recombination to suppress senescence in cells lacking telomerase.Proc Natl Acad Sci U S A. 2014 Mar 4;111(9):3377-82. doi: 10.1073/pnas.1307415111. Epub 2014 Feb 18. Proc Natl Acad Sci U S A. 2014. PMID: 24550456 Free PMC article.
-
Harmful R-loops are prevented via different cell cycle-specific mechanisms.Nat Commun. 2021 Jul 22;12(1):4451. doi: 10.1038/s41467-021-24737-x. Nat Commun. 2021. PMID: 34294712 Free PMC article.
-
Environmental stresses disrupt telomere length homeostasis.PLoS Genet. 2013;9(9):e1003721. doi: 10.1371/journal.pgen.1003721. Epub 2013 Sep 5. PLoS Genet. 2013. PMID: 24039592 Free PMC article.
-
The THO complex counteracts TERRA R-loop-mediated telomere fragility in telomerase+ cells and telomeric recombination in ALT+ cells.Nucleic Acids Res. 2023 Jul 21;51(13):6702-6722. doi: 10.1093/nar/gkad448. Nucleic Acids Res. 2023. PMID: 37246640 Free PMC article.
References
-
- Gilson E, Geli V. How telomeres are replicated. Nat Rev Mol Cell Biol. 2007;8:825–838. - PubMed
-
- Blackburn EH, Greider CW, Szostak JW. Telomeres and telomerase: the path from maize, Tetrahymena and yeast to human cancer and aging. Nat Med. 2006;12:1133–1138. - PubMed
-
- Collins K, Mitchell JR. Telomerase in the human organism. Oncogene. 2002;21:564–579. - PubMed
-
- Lingner J, Hughes TR, Shevchenko A, Mann M, Lundblad V, et al. Reverse transcriptase motifs on the catalytic subunit of telomerase. Science. 1997;276:561–567. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
