

Sodium channels and mammalian sensory mechanotransduction
- PMID: 22449024
- PMCID: PMC3378430
- DOI: 10.1186/1744-8069-8-21
Sodium channels and mammalian sensory mechanotransduction
Abstract
Background: Members of the degenerin/epithelial (DEG/ENaC) sodium channel family are mechanosensors in C elegans, and Nav1.7 and Nav1.8 voltage-gated sodium channel knockout mice have major deficits in mechanosensation. β and γENaC sodium channel subunits are present with acid sensing ion channels (ASICs) in mammalian sensory neurons of the dorsal root ganglia (DRG). The extent to which epithelial or voltage-gated sodium channels are involved in transduction of mechanical stimuli is unclear.
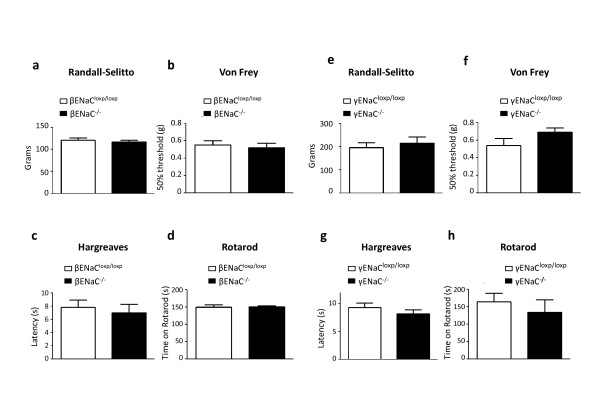
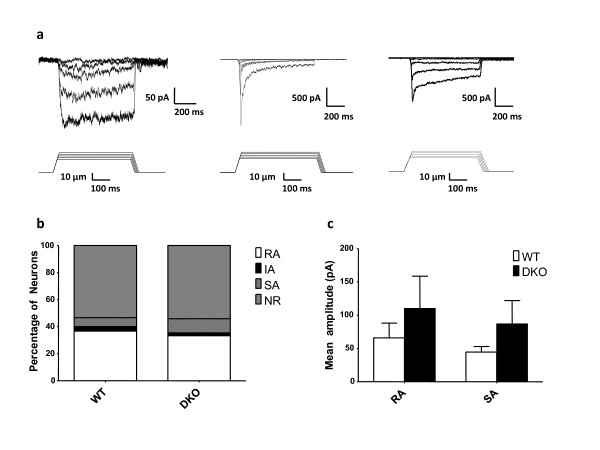
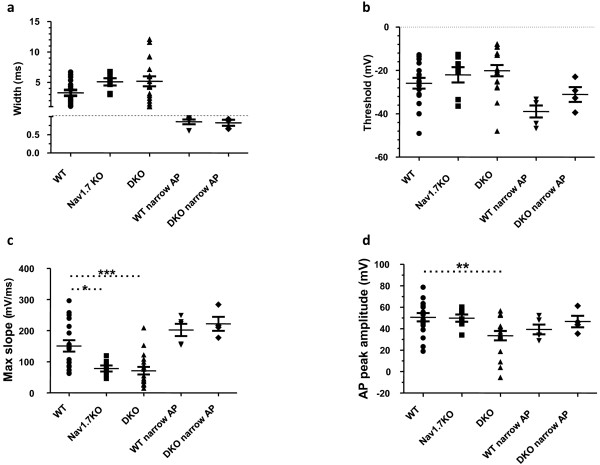
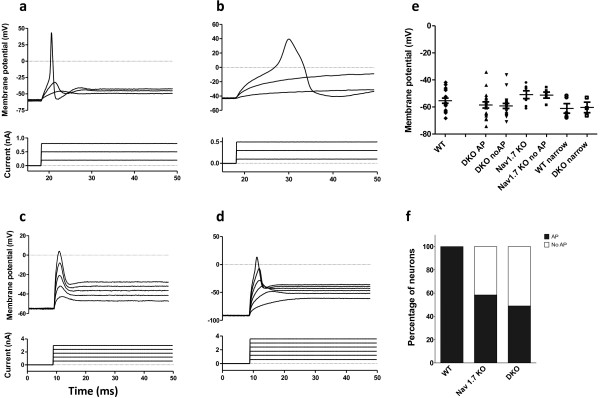
Results: Here we show that deleting β and γENaC sodium channels in sensory neurons does not result in mechanosensory behavioural deficits. We had shown previously that Nav1.7/Nav1.8 double knockout mice have major deficits in behavioural responses to noxious mechanical pressure. However, all classes of mechanically activated currents in DRG neurons are unaffected by deletion of the two sodium channels. In contrast, the ability of Nav1.7/Nav1.8 knockout DRG neurons to generate action potentials is compromised with 50% of the small diameter sensory neurons unable to respond to electrical stimulation in vitro.
Conclusion: Behavioural deficits in Nav1.7/Nav1.8 knockout mice reflects a failure of action potential propagation in a mechanosensitive set of sensory neurons rather than a loss of primary transduction currents. DEG/ENaC sodium channels are not mechanosensors in mouse sensory neurons.
Figures

References
-
- Delmas P, Hao J, Rodat-Despoix L. Molecular mechanisms of mechanotransduction in mammalian sensory neurons. Nat Rev Neurosci. 2011;12:139–153. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
