

The mechanistic basis for noncompetitive ibogaine inhibition of serotonin and dopamine transporters
- PMID: 22451652
- PMCID: PMC3365767
- DOI: 10.1074/jbc.M112.343681
The mechanistic basis for noncompetitive ibogaine inhibition of serotonin and dopamine transporters
Abstract
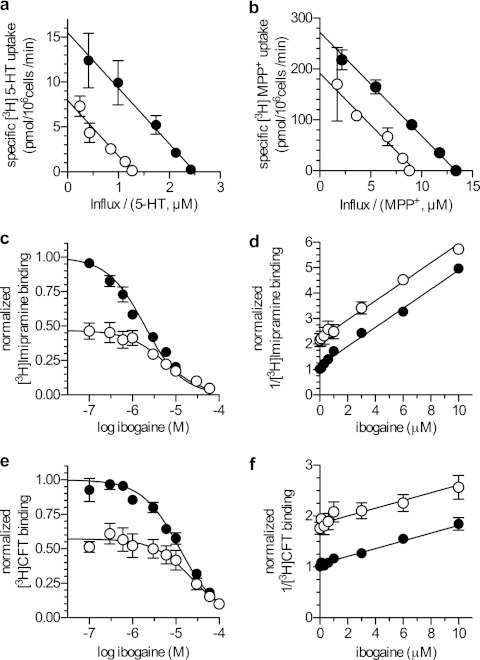
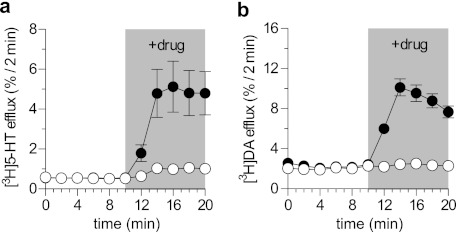
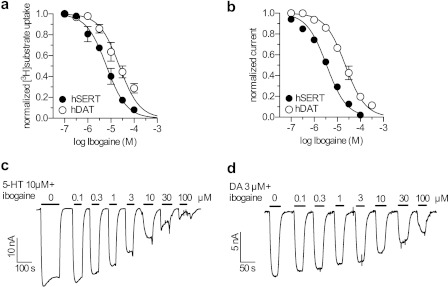
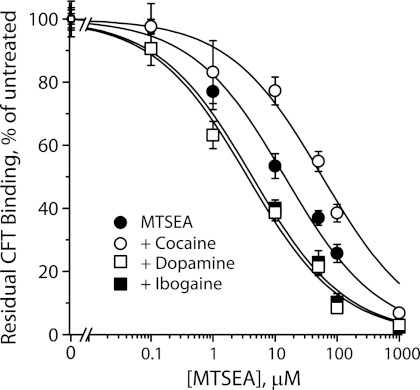
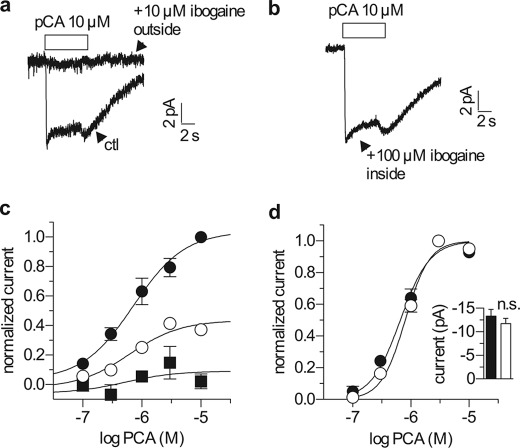
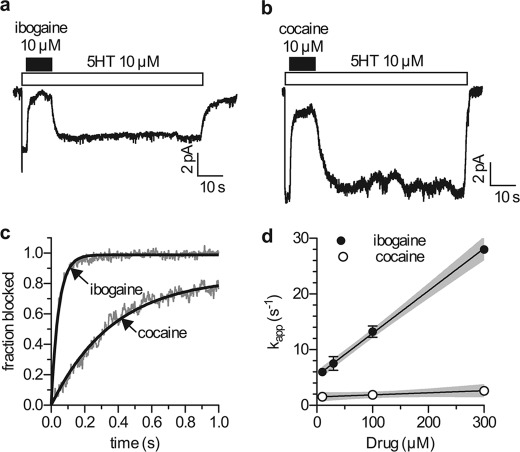
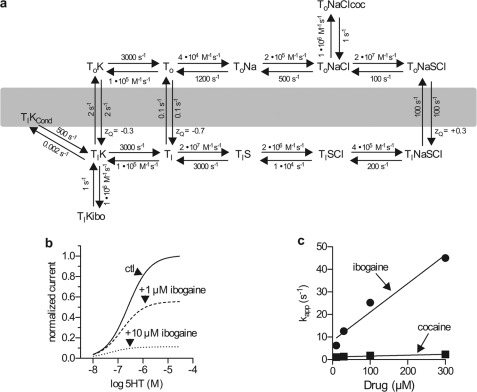
Ibogaine, a hallucinogenic alkaloid proposed as a treatment for opiate withdrawal, has been shown to inhibit serotonin transporter (SERT) noncompetitively, in contrast to all other known inhibitors, which are competitive with substrate. Ibogaine binding to SERT increases accessibility in the permeation pathway connecting the substrate-binding site with the cytoplasm. Because of the structural similarity between ibogaine and serotonin, it had been suggested that ibogaine binds to the substrate site of SERT. The results presented here show that ibogaine binds to a distinct site, accessible from the cell exterior, to inhibit both serotonin transport and serotonin-induced ionic currents. Ibogaine noncompetitively inhibited transport by both SERT and the homologous dopamine transporter (DAT). Ibogaine blocked substrate-induced currents also in DAT and increased accessibility of the DAT cytoplasmic permeation pathway. When present on the cell exterior, ibogaine inhibited SERT substrate-induced currents, but not when it was introduced into the cytoplasm through the patch electrode. Similar to noncompetitive transport inhibition, the current block was not reversed by increasing substrate concentration. The kinetics of inhibitor binding and dissociation, as determined by their effect on SERT currents, indicated that ibogaine does not inhibit by forming a long-lived complex with SERT, but rather binds directly to the transporter in an inward-open conformation. A kinetic model for transport describing the noncompetitive action of ibogaine and the competitive action of cocaine accounts well for the results of the present study.
Figures
References
-
- Leal M. B., Michelin K., Souza D. O., Elisabetsky E. (2003) Ibogaine attenuation of morphine withdrawal in mice. Role of glutamate N-methyl-d-aspartate receptors. Prog. Neuropsychopharmacol. Biol. Psychiatry 27, 781–785 - PubMed
-
- Levi M. S., Borne R. F. (2002) A review of chemical agents in the pharmacotherapy of addiction. Curr. Med. Chem. 9, 1807–1818 - PubMed
-
- Donnelly J. R. (2011) The need for ibogaine in drug and alcohol addiction treatment. J. Legal Med. 32, 93–114 - PubMed
-
- Jacobs M. T., Zhang Y. W., Campbell S. D., Rudnick G. (2007) Ibogaine, a noncompetitive inhibitor of serotonin transport, acts by stabilizing the cytoplasm-facing state of the transporter. J. Biol. Chem. 282, 29441–29447 - PubMed
-
- Wells G. B., Lopez M. C., Tanaka J. C. (1999) The effects of ibogaine on dopamine and serotonin transport in rat brain synaptosomes. Brain Res. Bull. 48, 641–647 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
