

The ATPase activity of MLH1 is required to orchestrate DNA double-strand breaks and end processing during class switch recombination
- PMID: 22451719
- PMCID: PMC3328365
- DOI: 10.1084/jem.20111531
The ATPase activity of MLH1 is required to orchestrate DNA double-strand breaks and end processing during class switch recombination
Abstract
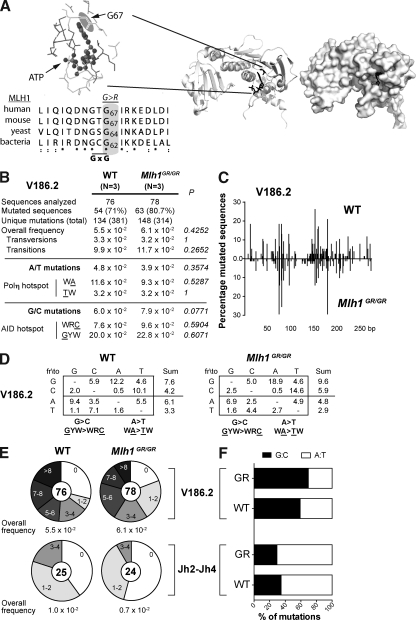
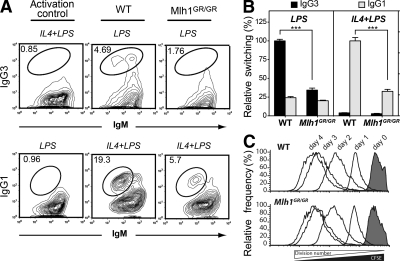
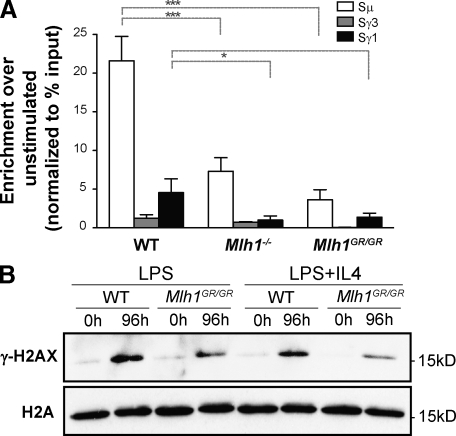
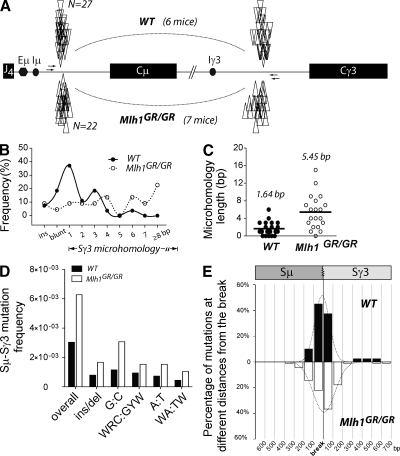
Antibody diversification through somatic hypermutation (SHM) and class switch recombination (CSR) are similarly initiated in B cells with the generation of U:G mismatches by activation-induced cytidine deaminase but differ in their subsequent mutagenic consequences. Although SHM relies on the generation of nondeleterious point mutations, CSR depends on the production of DNA double-strand breaks (DSBs) and their adequate recombination through nonhomologous end joining (NHEJ). MLH1, an ATPase member of the mismatch repair (MMR) machinery, is emerging as a likely regulator of whether a U:G mismatch progresses toward mutation or DSB formation. We conducted experiments on cancer modeled ATPase-deficient MLH1G67R knockin mice to determine the function that the ATPase domain of MLH1 mediates in SHM and CSR. Mlh1(GR/GR) mice displayed a significant decrease in CSR, mainly attributed to a reduction in the generation of DSBs and diminished accumulation of 53BP1 at the immunoglobulin switch regions. However, SHM was normal in these mice, which distinguishes MLH1 from upstream members of the MMR pathway and suggests a very specific role of its ATPase-dependent functions during CSR. In addition, we show that the residual switching events still taking place in Mlh1(GR/GR) mice display unique features, suggesting a role for the ATPase activity of MLH1 beyond the activation of the endonuclease functions of its MMR partner PMS2. A preference for switch junctions with longer microhomologies in Mlh1(GR/GR) mice suggests that through its ATPase activity, MLH1 also has an impact in DNA end processing, favoring canonical NHEJ downstream of the DSB. Collectively, our study shows that the ATPase domain of MLH1 is important to transmit the CSR signaling cascade both upstream and downstream of the generation of DSBs.
Figures
Similar articles
-
Mismatch repair proteins MSH2, MLH1, and EXO1 are important for class-switch recombination events occurring in B cells that lack nonhomologous end joining.J Immunol. 2011 Feb 15;186(4):2336-43. doi: 10.4049/jimmunol.1003104. Epub 2011 Jan 17. J Immunol. 2011. PMID: 21242524 Free PMC article.
-
Class switch recombination efficiency and junction microhomology patterns in Msh2-, Mlh1-, and Exo1-deficient mice depend on the presence of mu switch region tandem repeats.J Immunol. 2009 Jul 15;183(2):1222-8. doi: 10.4049/jimmunol.0900135. Epub 2009 Jun 24. J Immunol. 2009. PMID: 19553545 Free PMC article.
-
Fanca deficiency reduces A/T transitions in somatic hypermutation and alters class switch recombination junctions in mouse B cells.J Exp Med. 2014 Jun 2;211(6):1011-8. doi: 10.1084/jem.20131637. Epub 2014 May 5. J Exp Med. 2014. PMID: 24799500 Free PMC article.
-
Opinion: uracil DNA glycosylase (UNG) plays distinct and non-canonical roles in somatic hypermutation and class switch recombination.Int Immunol. 2014 Oct;26(10):575-8. doi: 10.1093/intimm/dxu071. Epub 2014 Jul 3. Int Immunol. 2014. PMID: 24994819 Free PMC article. Review.
-
Mechanism and control of V(D)J recombination versus class switch recombination: similarities and differences.Adv Immunol. 2005;86:43-112. doi: 10.1016/S0065-2776(04)86002-4. Adv Immunol. 2005. PMID: 15705419 Review.
Cited by
-
Downregulation of FOXP1 is required during germinal center B-cell function.Blood. 2013 May 23;121(21):4311-20. doi: 10.1182/blood-2012-10-462846. Epub 2013 Apr 11. Blood. 2013. PMID: 23580662 Free PMC article.
-
Dual Targeting of DNA Damage Response Proteins Implicated in Cancer Radioresistance.Genes (Basel). 2023 Dec 17;14(12):2227. doi: 10.3390/genes14122227. Genes (Basel). 2023. PMID: 38137049 Free PMC article.
-
The MLH1 ATPase domain is needed for suppressing aberrant formation of interstitial telomeric sequences.DNA Repair (Amst). 2018 May;65:20-25. doi: 10.1016/j.dnarep.2018.03.002. Epub 2018 Mar 7. DNA Repair (Amst). 2018. PMID: 29544212 Free PMC article.
-
Human MLH1 suppresses the insertion of telomeric sequences at intra-chromosomal sites in telomerase-expressing cells.Nucleic Acids Res. 2017 Feb 17;45(3):1219-1232. doi: 10.1093/nar/gkw1170. Nucleic Acids Res. 2017. PMID: 28180301 Free PMC article.
-
TDP43 Interacts with MLH1 and MSH6 Proteins in A DNA Damage-Inducible Manner.Res Sq [Preprint]. 2024 May 21:rs.3.rs-4439430. doi: 10.21203/rs.3.rs-4439430/v1. Res Sq. 2024. Update in: Mol Brain. 2024 Jun 5;17(1):32. doi: 10.1186/s13041-024-01108-3. PMID: 38826483 Free PMC article. Updated. Preprint.
References
-
- Avdievich E., Reiss C., Scherer S.J., Zhang Y., Maier S.M., Jin B., Hou H., Jr, Rosenwald A., Riedmiller H., Kucherlapati R., et al. 2008. Distinct effects of the recurrent Mlh1G67R mutation on MMR functions, cancer, and meiosis. Proc. Natl. Acad. Sci. USA. 105:4247–4252 10.1073/pnas.0800276105 - DOI - PMC - PubMed
-
- Bothmer A., Robbiani D.F., Di Virgilio M., Bunting S.F., Klein I.A., Feldhahn N., Barlow J., Chen H.T., Bosque D., Callen E., et al. 2011. Regulation of DNA end joining, resection, and immunoglobulin class switch recombination by 53BP1. Mol. Cell. 42:319–329 10.1016/j.molcel.2011.03.019 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
