

Conservation and divergence in Toll-like receptor 4-regulated gene expression in primary human versus mouse macrophages
- PMID: 22451944
- PMCID: PMC3341041
- DOI: 10.1073/pnas.1110156109
Conservation and divergence in Toll-like receptor 4-regulated gene expression in primary human versus mouse macrophages
Abstract
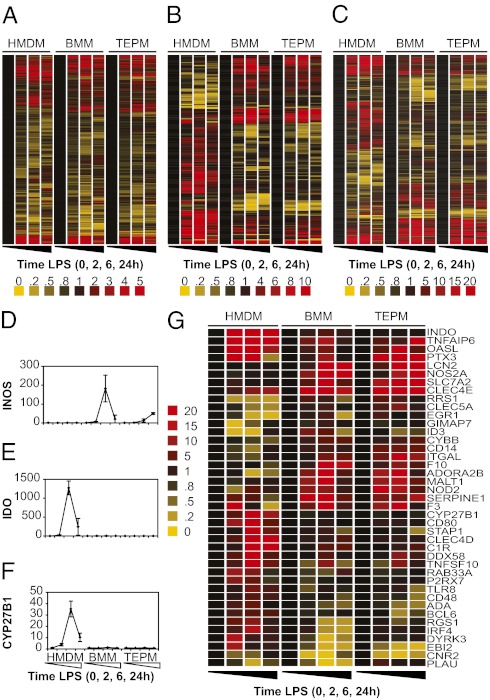
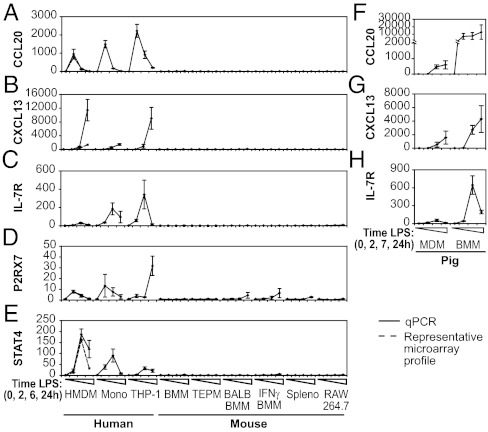
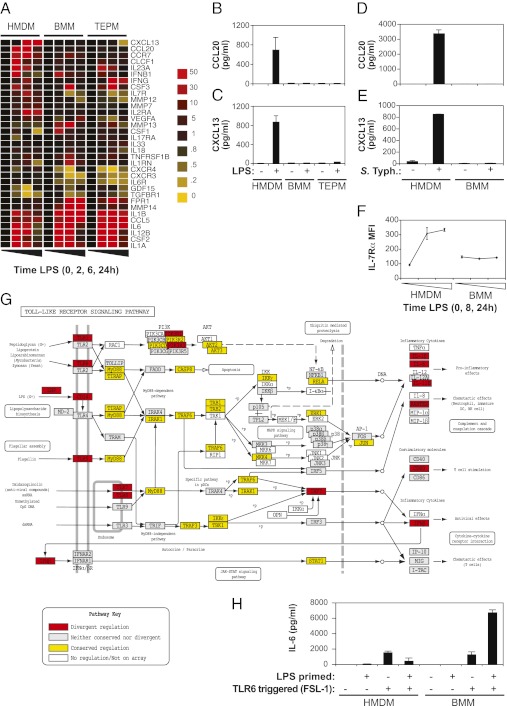
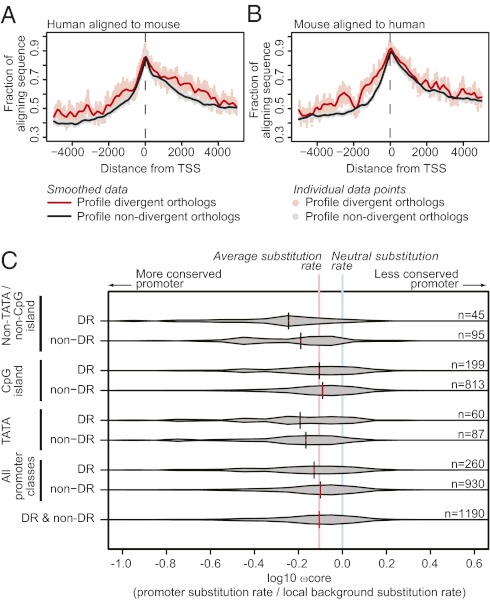
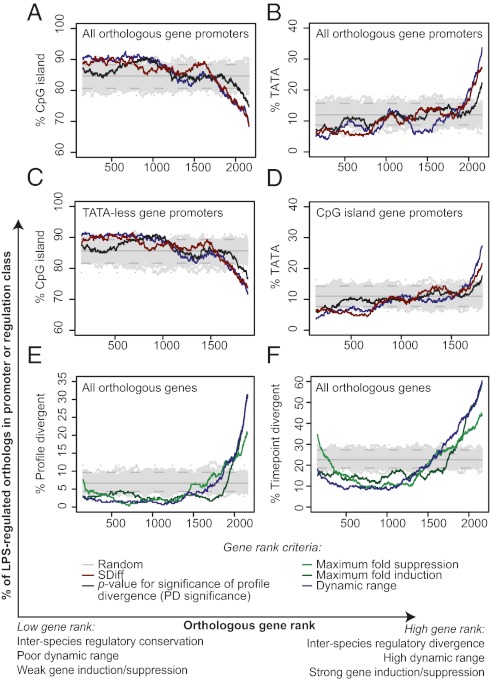
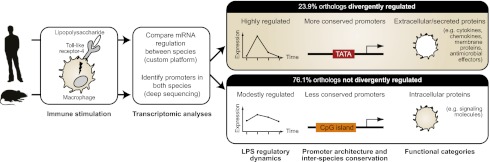
Evolutionary change in gene expression is generally considered to be a major driver of phenotypic differences between species. We investigated innate immune diversification by analyzing interspecies differences in the transcriptional responses of primary human and mouse macrophages to the Toll-like receptor (TLR)-4 agonist lipopolysaccharide (LPS). By using a custom platform permitting cross-species interrogation coupled with deep sequencing of mRNA 5' ends, we identified extensive divergence in LPS-regulated orthologous gene expression between humans and mice (24% of orthologues were identified as "divergently regulated"). We further demonstrate concordant regulation of human-specific LPS target genes in primary pig macrophages. Divergently regulated orthologues were enriched for genes encoding cellular "inputs" such as cell surface receptors (e.g., TLR6, IL-7Rα) and functional "outputs" such as inflammatory cytokines/chemokines (e.g., CCL20, CXCL13). Conversely, intracellular signaling components linking inputs to outputs were typically concordantly regulated. Functional consequences of divergent gene regulation were confirmed by showing LPS pretreatment boosts subsequent TLR6 responses in mouse but not human macrophages, in keeping with mouse-specific TLR6 induction. Divergently regulated genes were associated with a large dynamic range of gene expression, and specific promoter architectural features (TATA box enrichment, CpG island depletion). Surprisingly, regulatory divergence was also associated with enhanced interspecies promoter conservation. Thus, the genes controlled by complex, highly conserved promoters that facilitate dynamic regulation are also the most susceptible to evolutionary change.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Waterston RH, et al. Mouse Genome Sequencing Consortium Initial sequencing and comparative analysis of the mouse genome. Nature. 2002;420:520–562. - PubMed
-
- Mestas J, Hughes CC. Of mice and not men: Differences between mouse and human immunology. J Immunol. 2004;172:2731–2738. - PubMed
-
- Hayday AC, Peakman M. The habitual, diverse and surmountable obstacles to human immunology research. Nat Immunol. 2008;9:575–580. - PubMed
-
- Medzhitov R. Recognition of microorganisms and activation of the immune response. Nature. 2007;449:819–826. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
