

Gene transfer corrects acute GM2 gangliosidosis--potential therapeutic contribution of perivascular enzyme flow
- PMID: 22453766
- PMCID: PMC3412498
- DOI: 10.1038/mt.2012.44
Gene transfer corrects acute GM2 gangliosidosis--potential therapeutic contribution of perivascular enzyme flow
Abstract
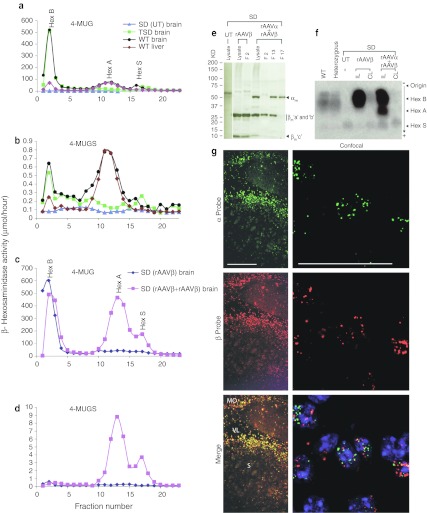
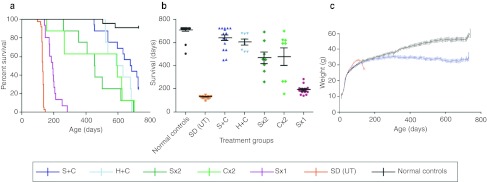
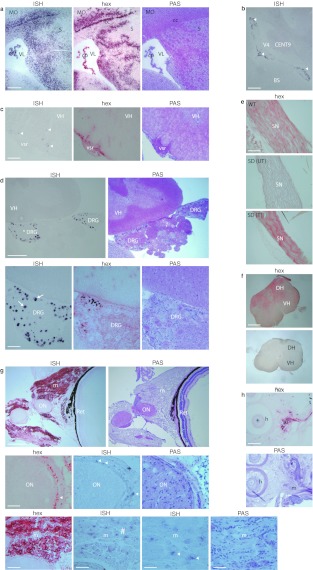
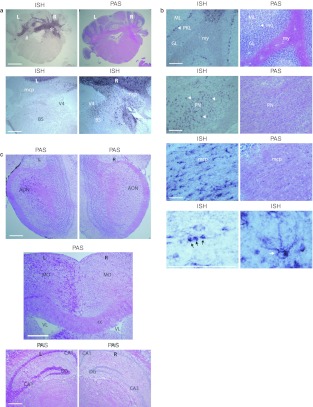
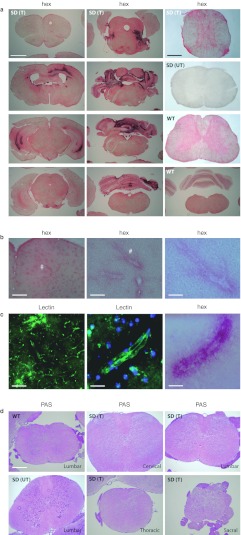
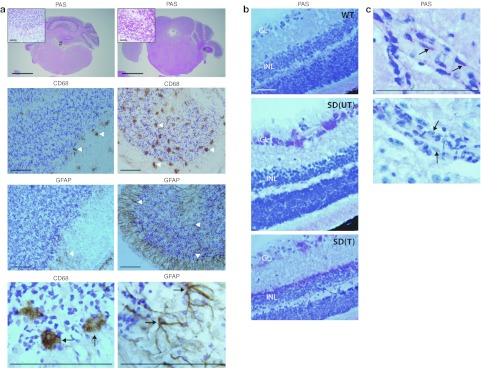
The GM2 gangliosidoses are fatal lysosomal storage diseases principally affecting the brain. Absence of β-hexosaminidase A and B activities in the Sandhoff mouse causes neurological dysfunction and recapitulates the acute Tay-Sachs (TSD) and Sandhoff diseases (SD) in infants. Intracranial coinjection of recombinant adeno-associated viral vectors (rAAV), serotype 2/1, expressing human β-hexosaminidase α (HEXA) and β (HEXB) subunits into 1-month-old Sandhoff mice gave unprecedented survival to 2 years and prevented disease throughout the brain and spinal cord. Classical manifestations of disease, including spasticity-as opposed to tremor-ataxia-were resolved by localized gene transfer to the striatum or cerebellum, respectively. Abundant biosynthesis of β-hexosaminidase isozymes and their global distribution via axonal, perivascular, and cerebrospinal fluid (CSF) spaces, as well as diffusion, account for the sustained phenotypic rescue-long-term protein expression by transduced brain parenchyma, choroid plexus epithelium, and dorsal root ganglia neurons supplies the corrective enzyme. Prolonged survival permitted expression of cryptic disease in organs not accessed by intracranial vector delivery. We contend that infusion of rAAV into CSF space and intraparenchymal administration by convection-enhanced delivery at a few strategic sites will optimally treat neurodegeneration in many diseases affecting the nervous system.
Figures
References
-
- Johnson WG. The clinical spectrum of hexosaminidase deficiency diseases. Neurology. 1981;31:1453–1456. - PubMed
-
- Kolodny EH, Brady RO., and, Volk BW. Demonstration of an alteration of ganglioside metabolism in Tay-Sachs disease. Biochem Biophys Res Commun. 1969;37:526–531. - PubMed
-
- Okada S., and, O'Brien JS. Tay-Sachs disease: generalized absence of a beta-D-N-acetylhexosaminidase component. Science. 1969;165:698–700. - PubMed
-
- Sandhoff K. Variation of beta-N-acetylhexosaminidase-pattern in Tay-Sachs disease. FEBS Lett. 1969;4:351–354. - PubMed
-
- Conzelmann E., and, Sandhoff K. Purification and characterization of an activator protein for the degradation of glycolipids GM2 and GA2 by hexosaminidase A. Hoppe-Seyler's Z Physiol Chem. 1979;360:1837–1849. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
