

Proteomic cornerstones of hematopoietic stem cell differentiation: distinct signatures of multipotent progenitors and myeloid committed cells
- PMID: 22454540
- PMCID: PMC3412962
- DOI: 10.1074/mcp.M111.016790
Proteomic cornerstones of hematopoietic stem cell differentiation: distinct signatures of multipotent progenitors and myeloid committed cells
Abstract
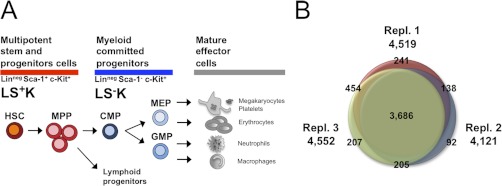
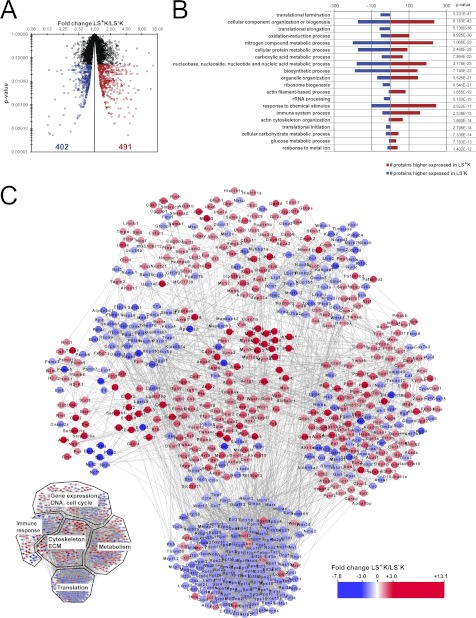
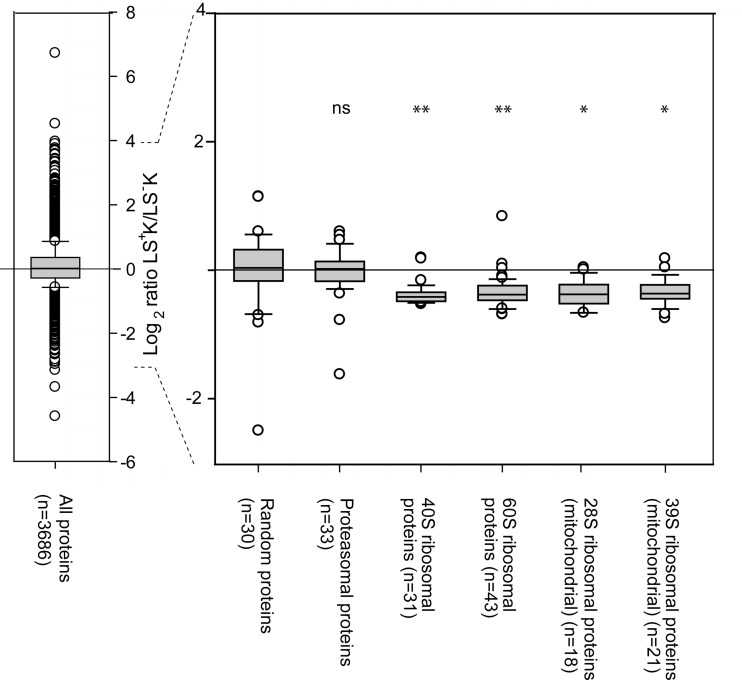
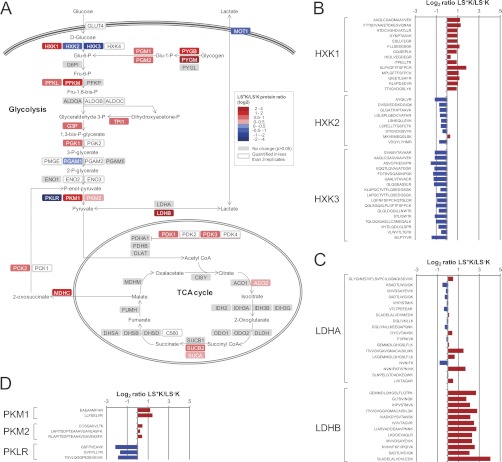
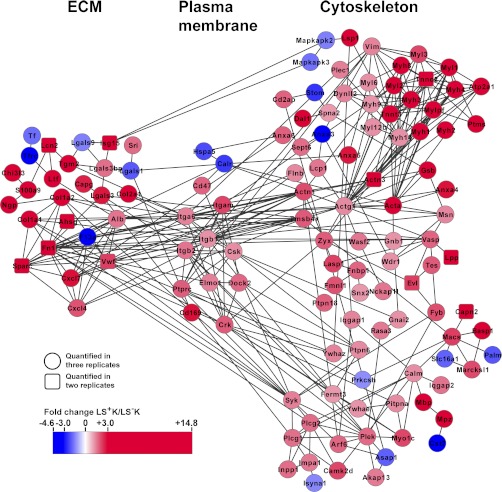
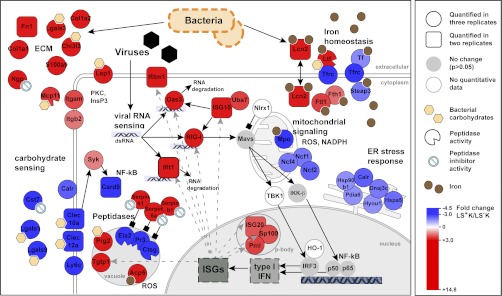
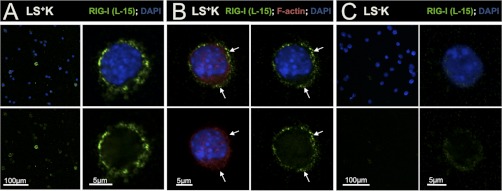
Regenerative tissues such as the skin epidermis, the intestinal mucosa or the hematopoietic system are organized in a hierarchical manner with stem cells building the top of this hierarchy. Somatic stem cells harbor the highest self-renewal activity and generate a series of multipotent progenitors which differentiate into lineage committed progenitors and subsequently mature cells. In this report, we applied an in-depth quantitative proteomic approach to analyze and compare the full proteomes of ex vivo isolated and FACS-sorted populations highly enriched for either multipotent hematopoietic stem/progenitor cells (HSPCs, Lin(neg)Sca-1(+)c-Kit(+)) or myeloid committed precursors (Lin(neg)Sca-1(-)c-Kit(+)). By employing stable isotope dimethyl labeling and high-resolution mass spectrometry, more than 5000 proteins were quantified. From biological triplicate experiments subjected to rigorous statistical evaluation, 893 proteins were found differentially expressed between multipotent and myeloid committed cells. The differential protein content in these cell populations points to a distinct structural organization of the cytoskeleton including remodeling activity. In addition, we found a marked difference in the expression of metabolic enzymes, including a clear shift of specific protein isoforms of the glycolytic pathway. Proteins involved in translation showed a collective higher expression in myeloid progenitors, indicating an increased translational activity. Strikingly, the data uncover a unique signature related to immune defense mechanisms, centering on the RIG-I and type-1 interferon response systems, which are installed in multipotent progenitors but not evident in myeloid committed cells. This suggests that specific, and so far unrecognized, mechanisms protect these immature cells before they mature. In conclusion, this study indicates that the transition of hematopoietic stem/progenitors toward myeloid commitment is accompanied by a profound change in processing of cellular resources, adding novel insights into the molecular mechanisms at the interface between multipotency and lineage commitment.
Figures
References
-
- Till J. E., McCulloch C. E. (1961) A direct measurement of the radiation sensitivity of normal mouse bone marrow cells. Radiation Res. 14, 213–222 - PubMed
-
- Purton L. E., Scadden D. T. (2007) Limiting factors in murine hematopoietic stem cell assays. Cell Stem Cell 1, 263–270 - PubMed
-
- Wilson A., Oser G. M., Jaworski M., Blanco-Bose W. E., Laurenti E., Adolphe C., Essers M. A., Macdonald H. R., Trumpp A. (2007) Dormant and self-renewing hematopoietic stem cells and their niches. Ann. NY Acad. Sci. 1106, 64–75 - PubMed
-
- Trumpp A., Essers M., Wilson A. (2010) Awakening dormant haematopoietic stem cells. Nature Rev. Immunol. 10, 201–209 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
