

Fluorescence-based incision assay for human XPF-ERCC1 activity identifies important elements of DNA junction recognition
- PMID: 22457069
- PMCID: PMC3401468
- DOI: 10.1093/nar/gks284
Fluorescence-based incision assay for human XPF-ERCC1 activity identifies important elements of DNA junction recognition
Abstract
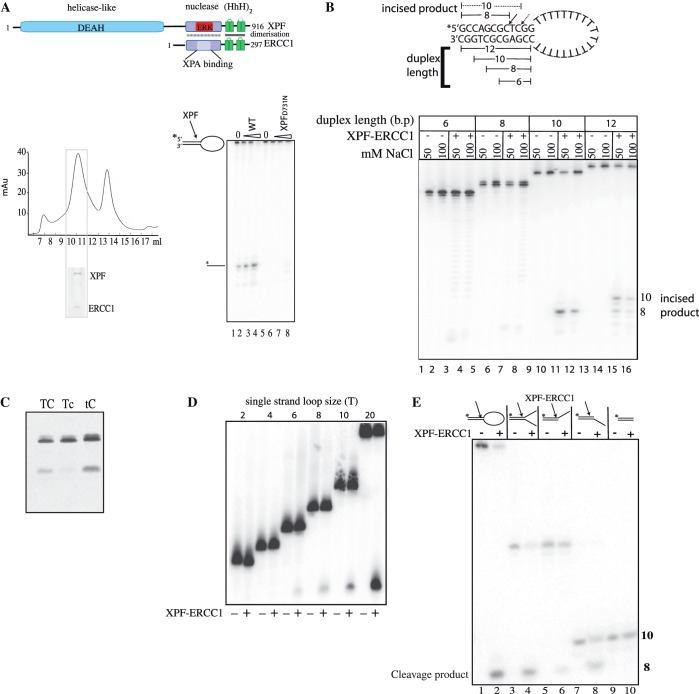
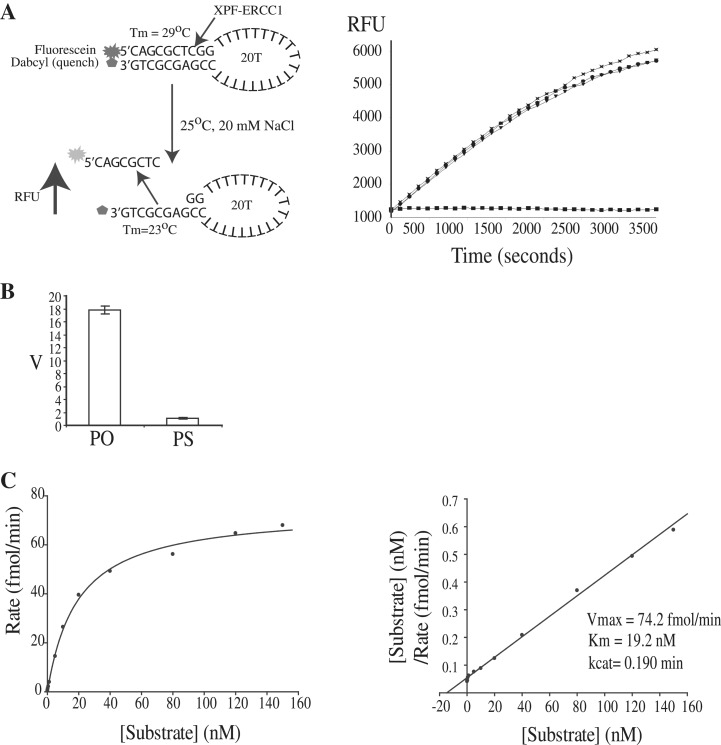
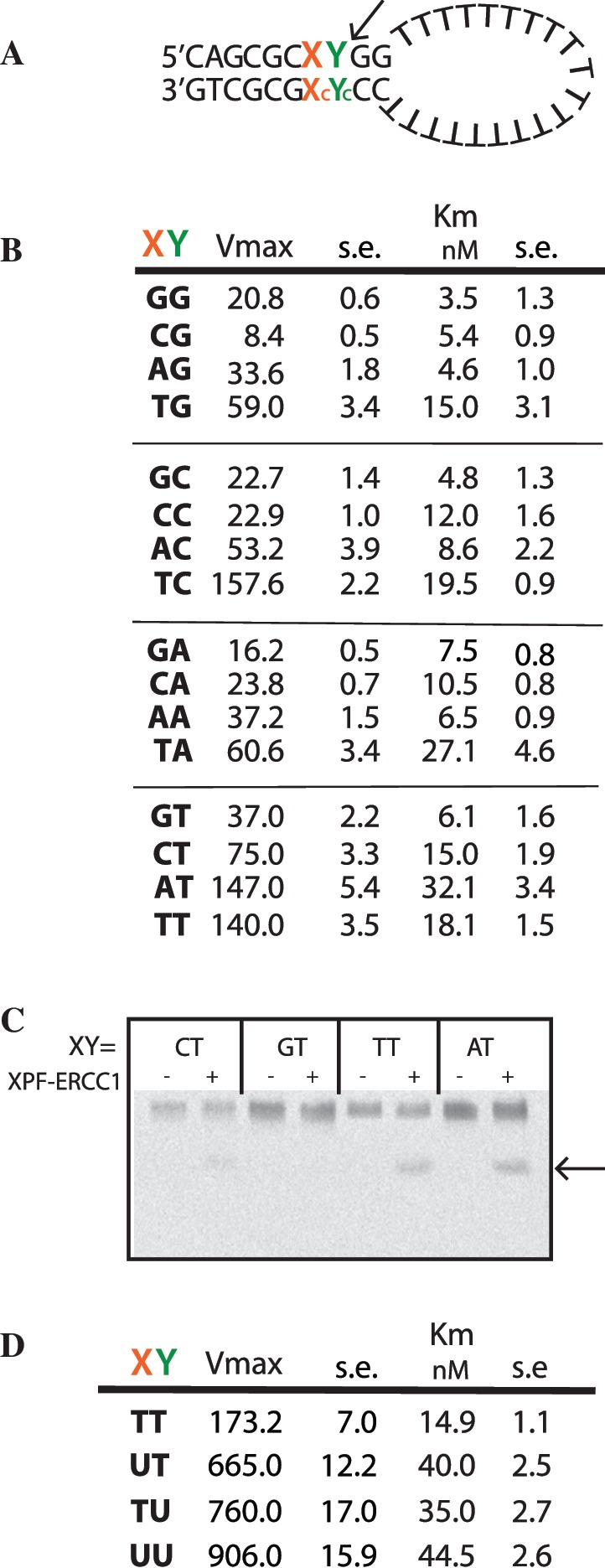
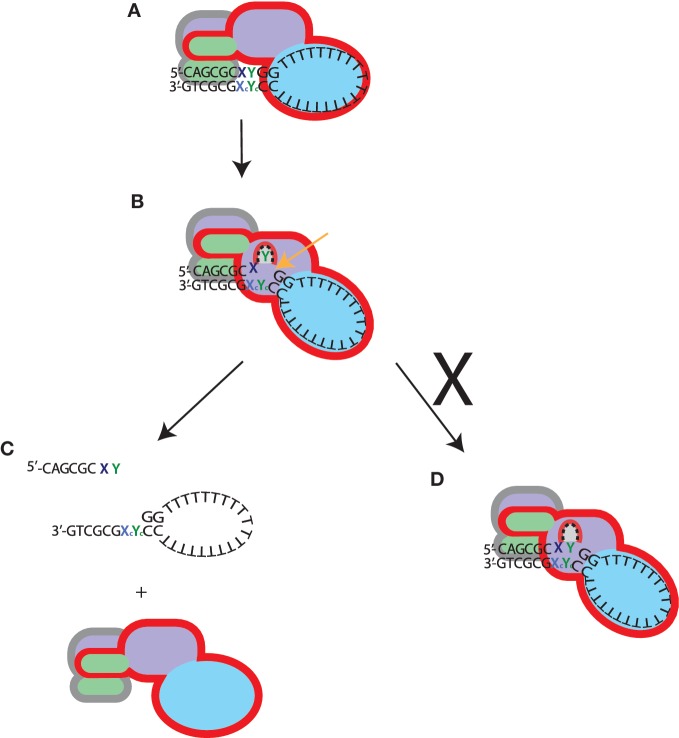
The structure-specific endonuclease activity of the human XPF-ERCC1 complex is essential for a number of DNA processing mechanisms that help to maintain genomic integrity. XPF-ERCC1 cleaves DNA structures such as stem-loops, bubbles or flaps in one strand of a duplex where there is at least one downstream single strand. Here, we define the minimal substrate requirements for cleavage of stem-loop substrates allowing us to develop a real-time fluorescence-based assay to measure endonuclease activity. Using this assay, we show that changes in the sequence of the duplex upstream of the incision site results in up to 100-fold variation in cleavage rate of a stem-loop substrate by XPF-ERCC1. XPF-ERCC1 has a preference for cleaving the phosphodiester bond positioned on the 3'-side of a T or a U, which is flanked by an upstream T or U suggesting that a T/U pocket may exist within the catalytic domain. In addition to an endonuclease domain and tandem helix-hairpin-helix domains, XPF has a divergent and inactive DEAH helicase-like domain (HLD). We show that deletion of HLD eliminates endonuclease activity and demonstrate that purified recombinant XPF-HLD shows a preference for binding stem-loop structures over single strand or duplex alone, suggesting a role for the HLD in initial structure recognition. Together our data describe features of XPF-ERCC1 and an accepted model substrate that are important for recognition and efficient incision activity.
Figures

References
-
- Friedberg EC, Aguilera A, Gellert M, Hanawalt PC, Hays JB, Lehmann AR, Lindahl T, Lowndes N, Sarasin A, Wood RD. DNA repair: from molecular mechanism to human disease. DNA Repair. 2006;5:986–996. - PubMed
-
- Sijbers AM, de Laat WL, Ariza RR, Biggerstaff M, Wei YF, Moggs JG, Carter KC, Shell BK, Evans E, de Jong MC, et al. Xeroderma pigmentosum group F caused by a defect in a structure-specific DNA repair endonuclease. Cell. 1996;86:811–822. - PubMed
-
- Niedernhofer LJ, Odijk H, Budzowska M, van Drunen E, Maas A, Theil AF, de Wit J, Jaspers NG, Beverloo HB, Hoeijmakers JH, et al. The structure-specific endonuclease Ercc1-Xpf is required to resolve DNA interstrand cross-link-induced double-strand breaks. Mol. Cell Biol. 2004;24:5776–5787. - PMC - PubMed
