

Editing T cell specificity towards leukemia by zinc finger nucleases and lentiviral gene transfer
- PMID: 22466705
- PMCID: PMC5019824
- DOI: 10.1038/nm.2700
Editing T cell specificity towards leukemia by zinc finger nucleases and lentiviral gene transfer
Abstract
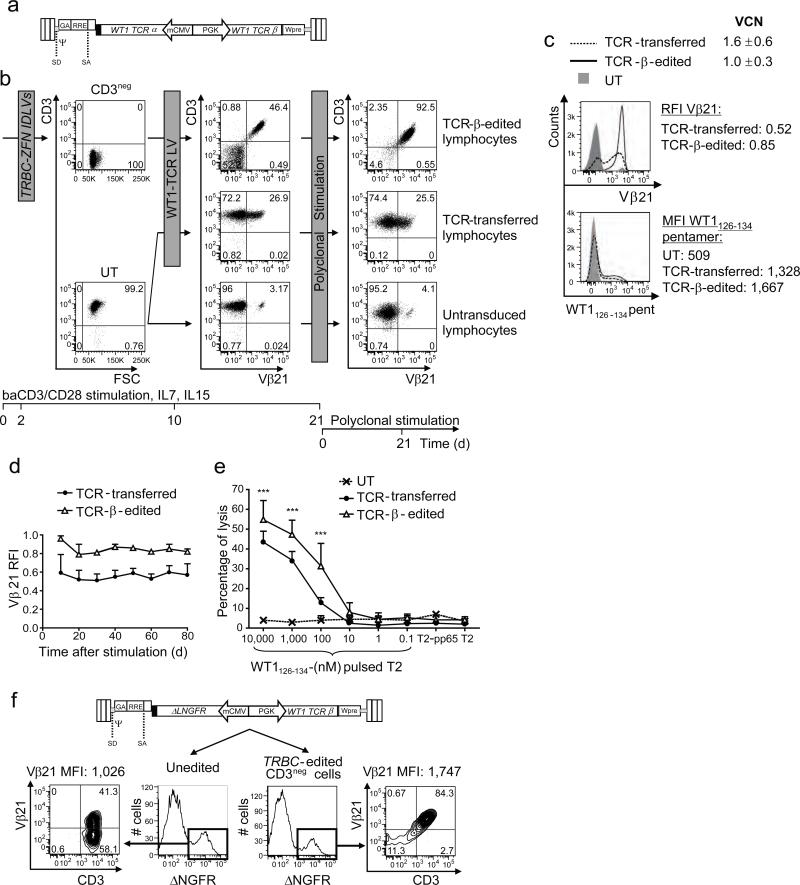
The transfer of high-avidity T cell receptor (TCR) genes isolated from rare tumor-specific lymphocytes into polyclonal T cells is an attractive cancer immunotherapy strategy. However, TCR gene transfer results in competition for surface expression and inappropriate pairing between the exogenous and endogenous TCR chains, resulting in suboptimal activity and potentially harmful unpredicted antigen specificities of the resultant TCRs. We designed zinc-finger nucleases (ZFNs) that promoted the disruption of endogenous TCR β- and α-chain genes. Lymphocytes treated with ZFNs lacked surface expression of CD3-TCR and expanded with the addition of interleukin-7 (IL-7) and IL-15. After lentiviral transfer of a TCR specific for the Wilms tumor 1 (WT1) antigen, these TCR-edited cells expressed the new TCR at high levels, were easily expanded to near purity and were superior at specific antigen recognition compared to donor-matched, unedited TCR-transferred cells. In contrast to unedited TCR-transferred cells, the TCR-edited lymphocytes did not mediate off-target reactivity while maintaining their anti-tumor activity in vivo, thus showing that complete editing of T cell specificity generates tumor-specific lymphocytes with improved biosafety profiles.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
