

Characterization of the Group A Streptococcus Mga virulence regulator reveals a role for the C-terminal region in oligomerization and transcriptional activation
- PMID: 22468267
- PMCID: PMC3324284
- DOI: 10.1111/j.1365-2958.2012.07980.x
Characterization of the Group A Streptococcus Mga virulence regulator reveals a role for the C-terminal region in oligomerization and transcriptional activation
Abstract
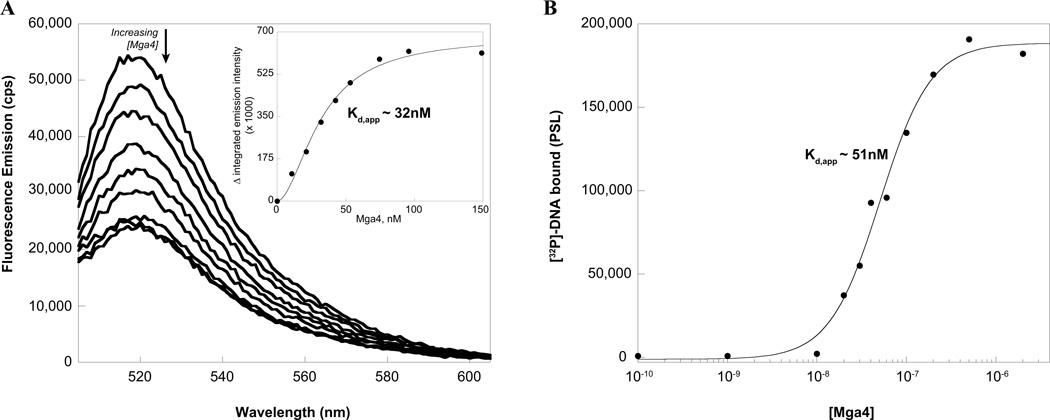
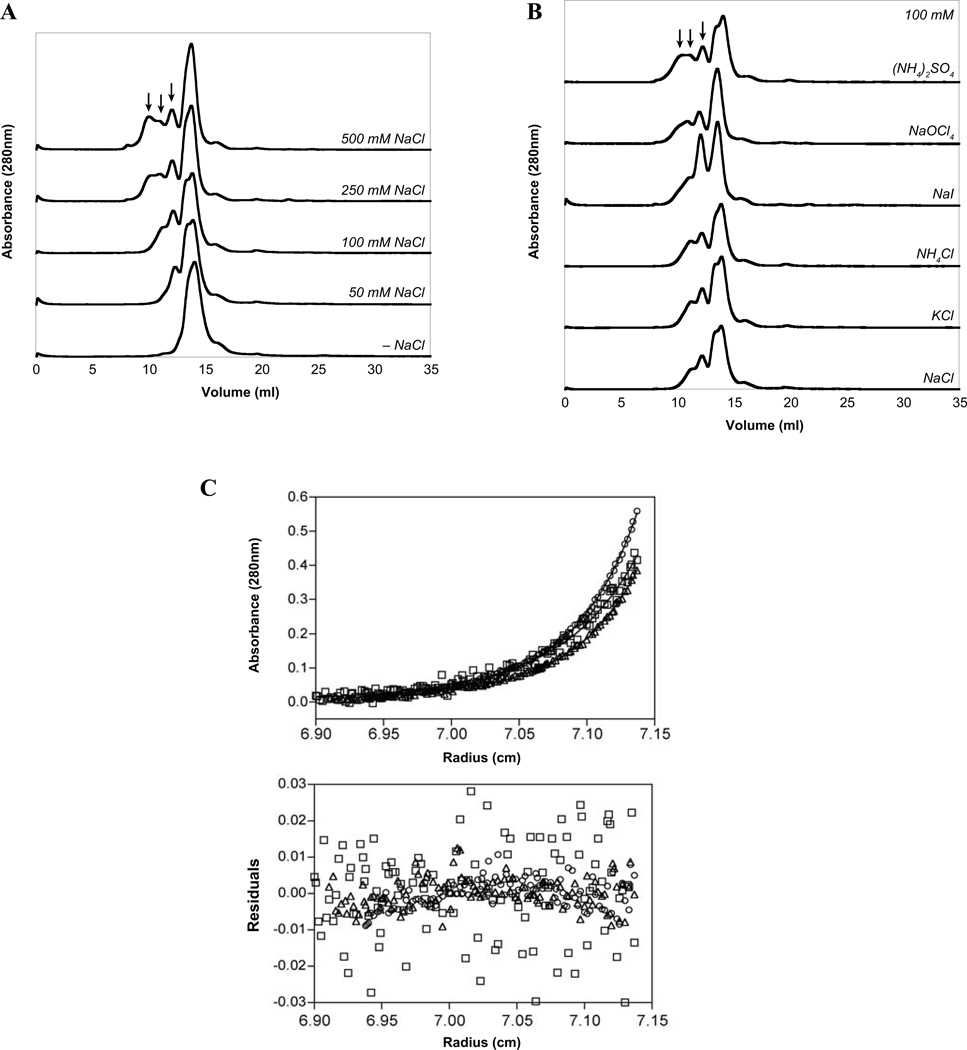
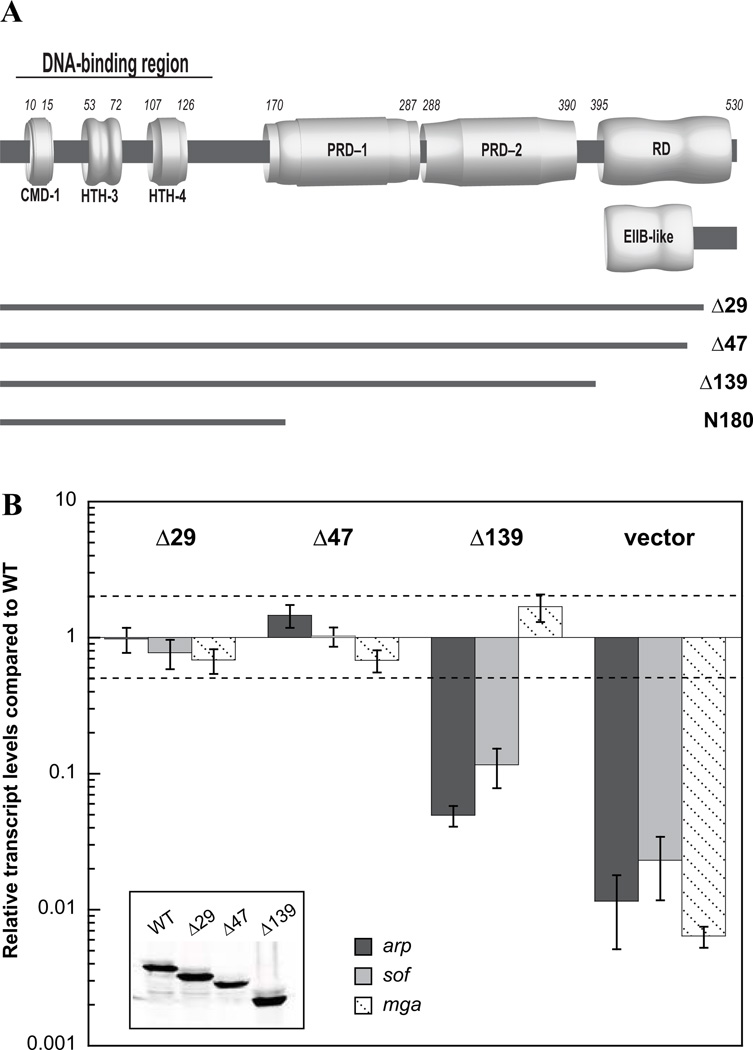
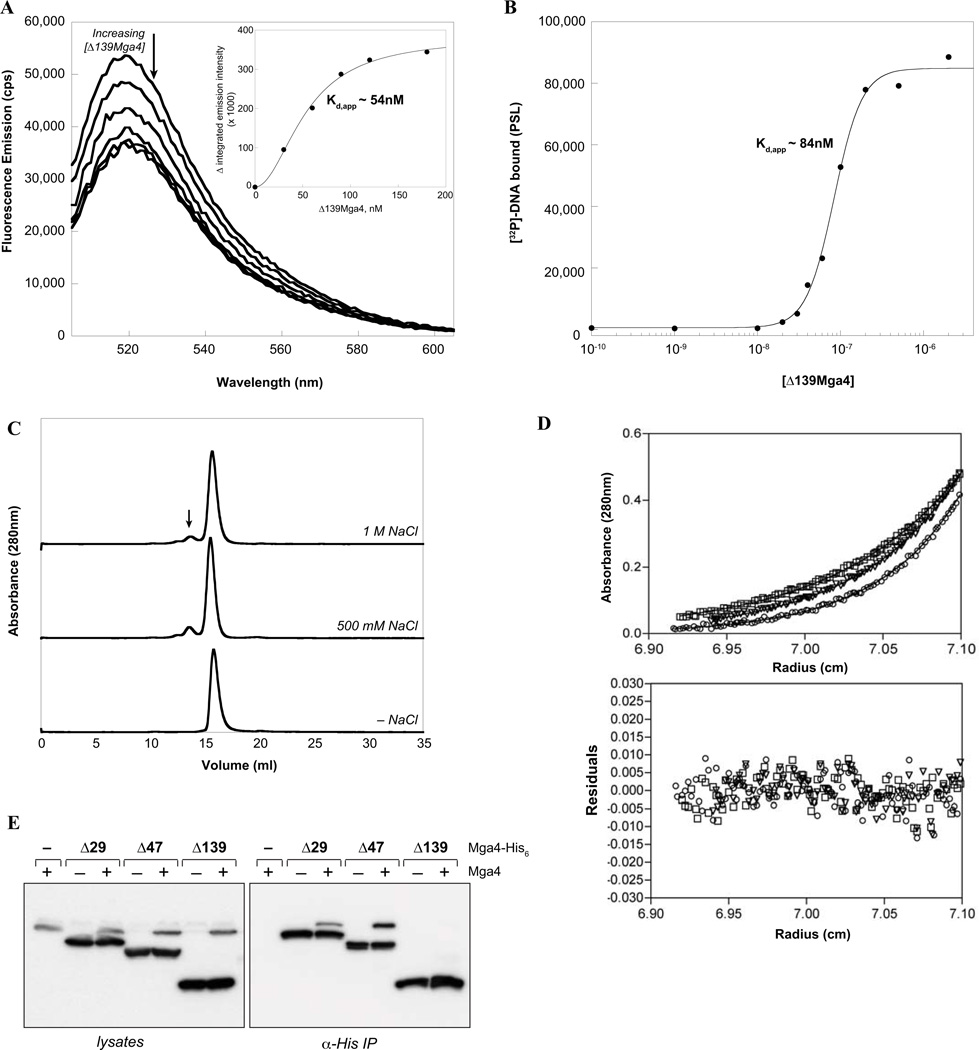
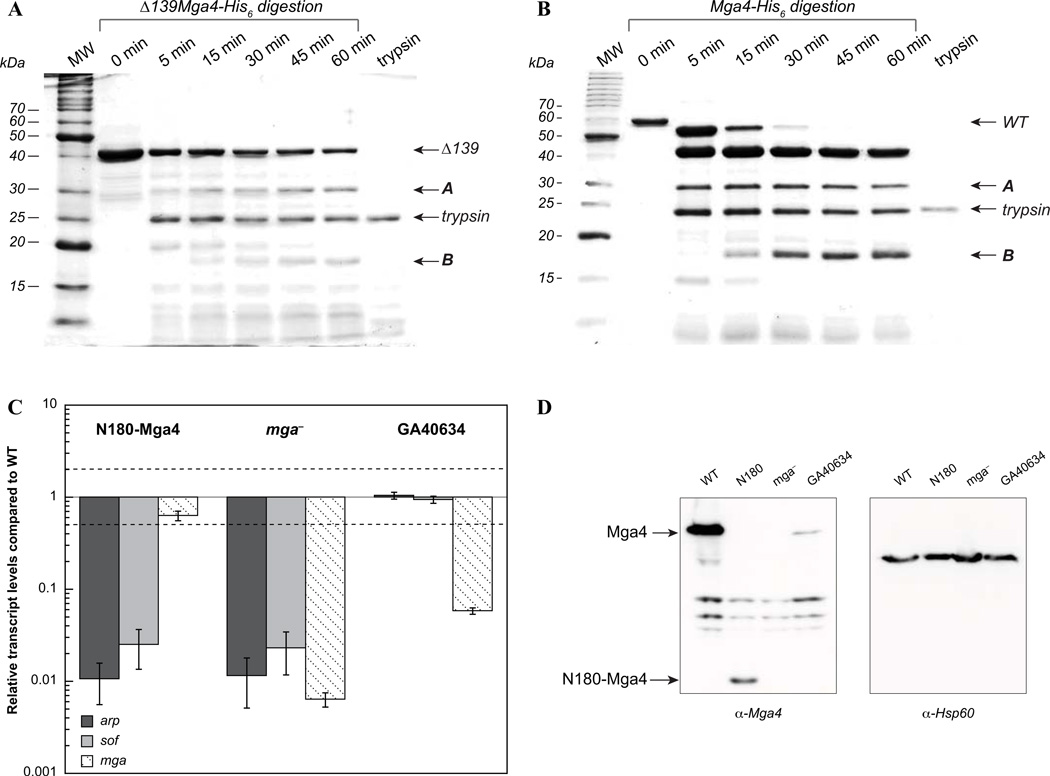
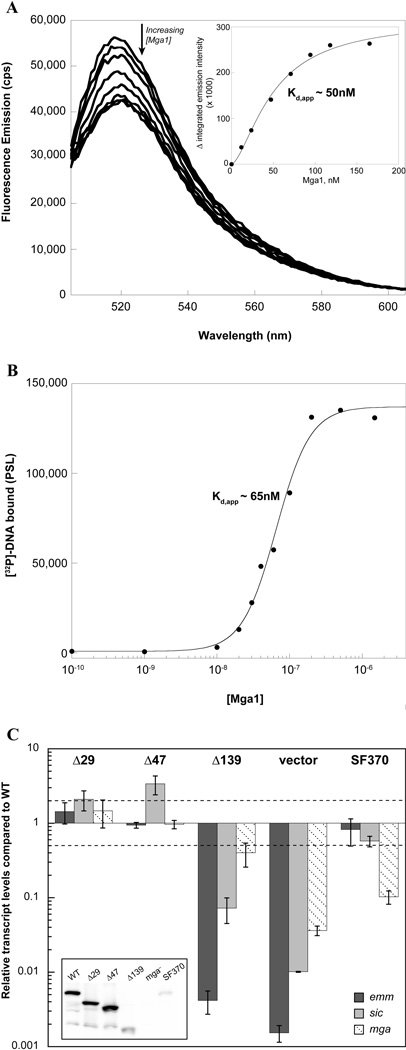
The Group A Streptococcus (GAS) is a strict human pathogen that causes a broad spectrum of illnesses. One of the key regulators of virulence in GAS is the transcriptional activator Mga, which co-ordinates the early stages of infection. Although the targets of Mga have been well characterized, basic biochemical analyses have been limited due to difficulties in obtaining purified protein. In this study, high-level purification of soluble Mga was achieved, enabling the first detailed characterization of the protein. Fluorescence titrations coupled with filter-binding assays indicate that Mga binds cognate DNA with nanomolar affinity. Gel filtration analyses, analytical ultracentrifugation and co-immunoprecipitation experiments demonstrate that Mga forms oligomers in solution.Moreover, the ability of the protein to oligomerize in solution was found to correlate with transcriptional activation; DNA binding appears to be necessary but insufficient for full activity. Truncation analyses reveal that the uncharacterized C-terminal region of Mga, possessing similarity to phosphotransferase system EIIB proteins, plays a critical role in oligomerization and in vivo activity. Mga from a divergent serotype was found to behave similarly, suggesting that this study describes a general mechanism for Mga regulation of target virulence genes within GAS and provides insight into related regulators in other Gram-positive pathogens.
Figures
References
-
- Andersson G, McIver K, Heden LO, Scott JR. Complementation of divergent mga genes in group A streptococcus. Gene. 1996;175:77–81. - PubMed
-
- Aravind L, Anantharaman V, Balaji S, Babu MM, Iyer LM. The many faces of the helix-turn-helix domain: transcription regulation and beyond. FEMS Microbiol Rev. 2005;29:231–262. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
