

Aryl hydrocarbon receptor ligand effects in RBL2H3 cells
- PMID: 22471748
- PMCID: PMC3529589
- DOI: 10.3109/1547691X.2012.661802
Aryl hydrocarbon receptor ligand effects in RBL2H3 cells
Abstract
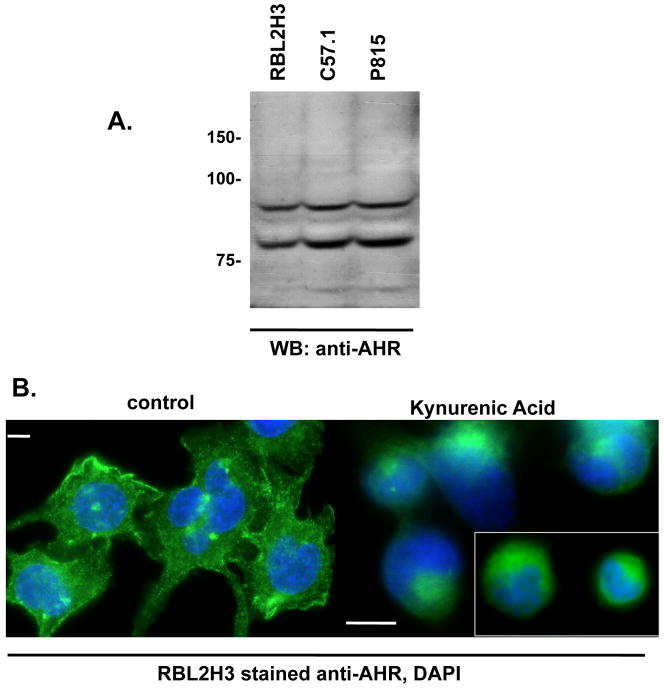
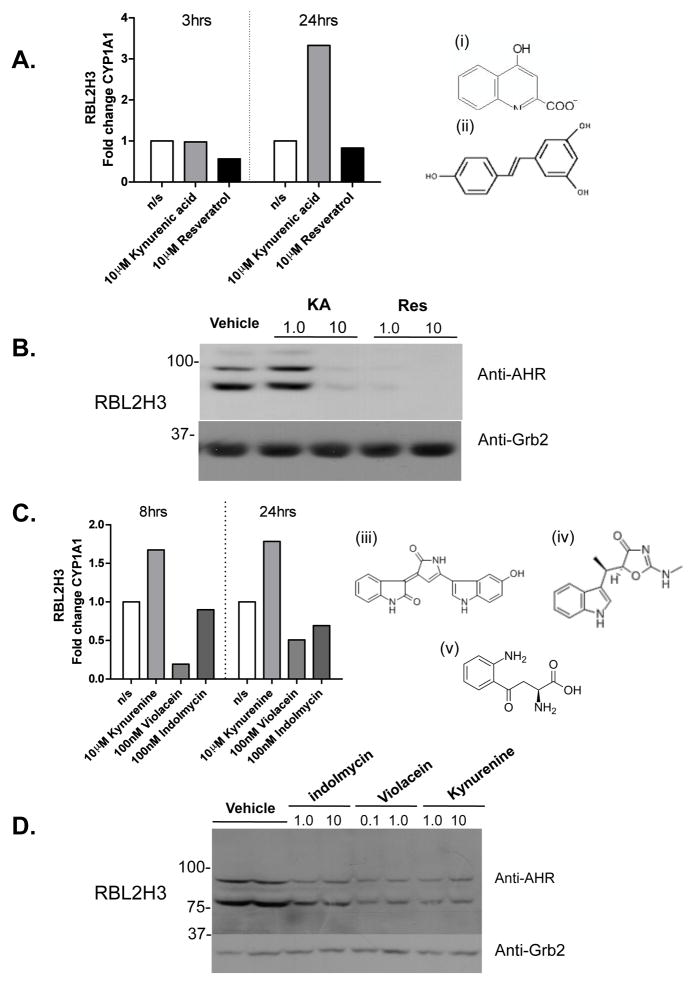
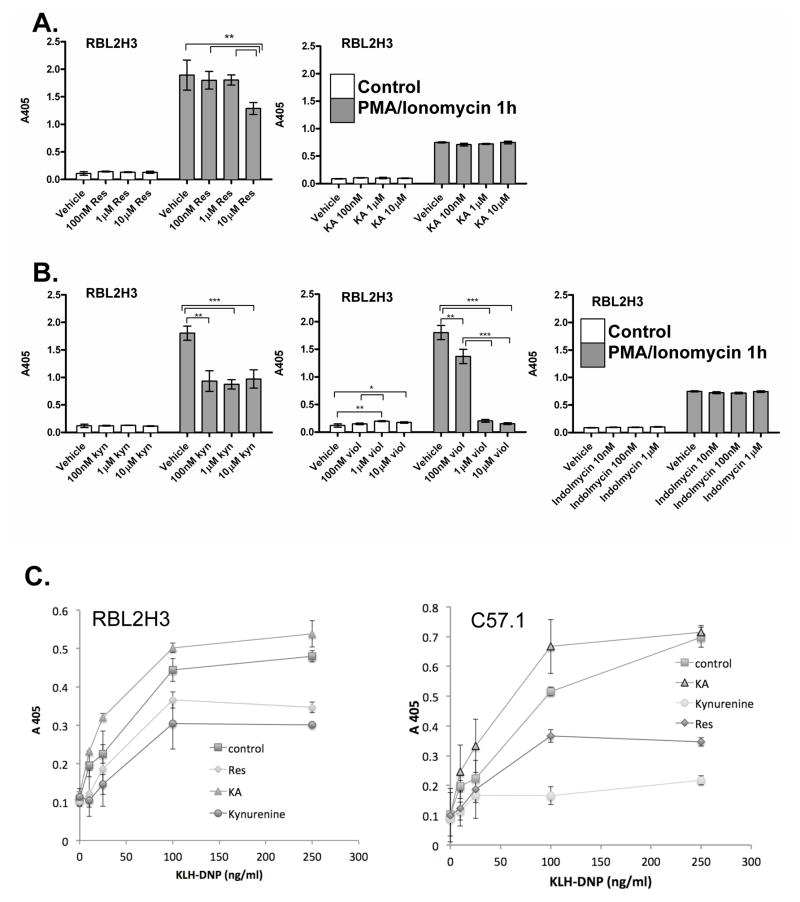
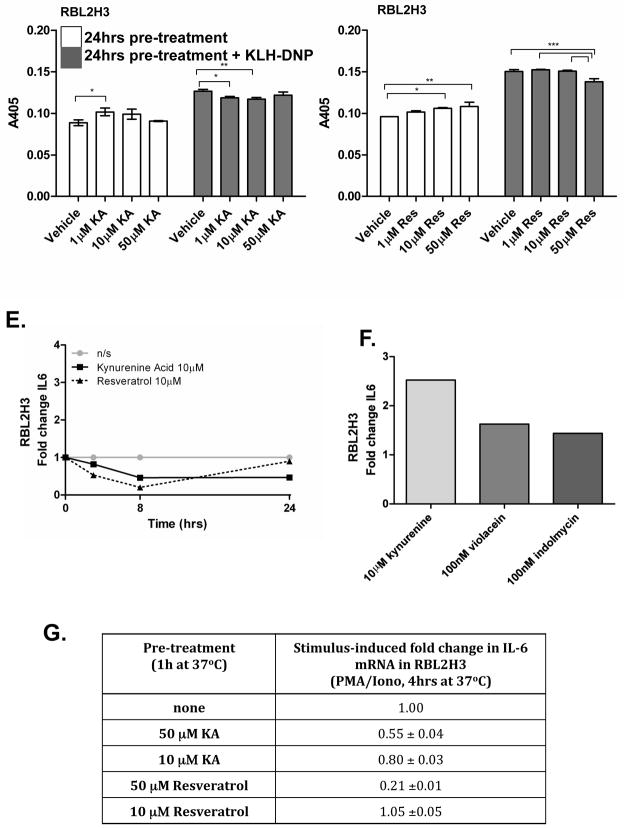
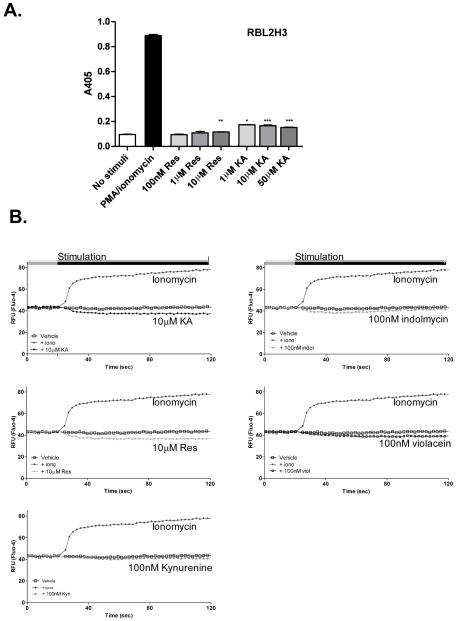
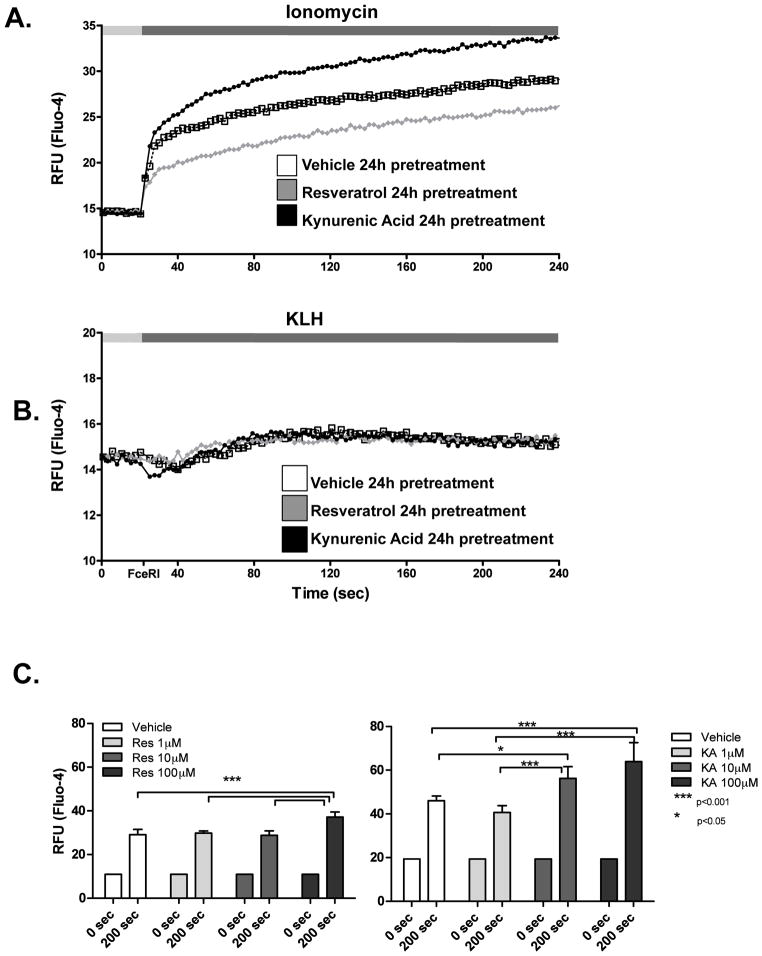
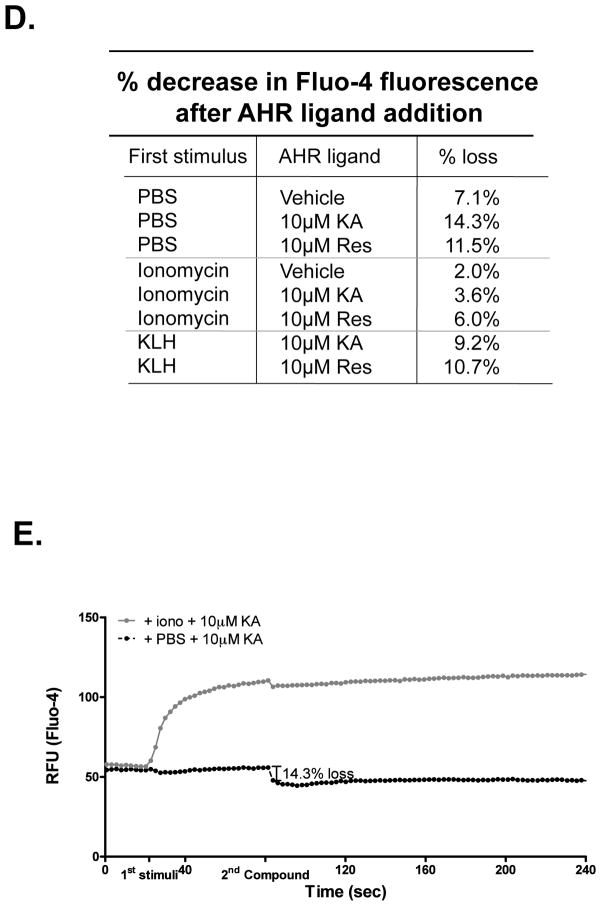
The aryl hydrocarbon receptor (AHR) mediates toxic effects of dioxin and xenobiotic metabolism. AHR has an emerging role in the immune system, but its physiological ligands and functional role in immunocytes remain poorly understood. Mast cells are immunocytes that are central to inflammatory responses and release a spectrum of pro-inflammatory mediators including histamine, mast cell proteases, and pro-inflammatory cytokines such as IL-6 upon stimulation. The aim was to investigate the AHR in model mast cells and examine how both putative and known AHR ligands, e.g., kynurenine, kynurenic acid (KA), Resveratrol, indolmycin, and violacein, affect mast cell activation and signaling. These ligands were tested on calcium signaling, degranulation, and gene expression. The data show that AHR is present in three model mast cell lines, and that various known and putative AHR ligands regulate gene expression of Cyp1a1, a gene down-stream of AHR. Furthermore, it was found that calcium influxes and mast cell secretory responses were enhanced or suppressed after chronic treatment with AHR agonists or antagonists, and that AHR ligands modified RBL2H3 cell degranulation. AHR ligands can chronically change cytokine gene expression in activated mast cells, as exemplified by IL-6. The antagonist Resveratrol repressed expression of induced IL-6 gene expression. Although KA and kynurenine are both AHR agonists, these ligands behaved differently in regards to degranulation and IL-6 expression, indicating that they may function outside of AHR pathways. These data suggest considerable complexity in RBL2H3 responses to AHR ligands, with implications for understanding of both dioxin pathology and the immunological effects of endogenous AHR ligands.
Conflict of interest statement
The authors report no declaration of interest. The authors alone are responsible for the content of this manuscript.
Figures
References
-
- Adachi J, Mori Y, Matsui S, Matsuda T. Comparison of gene expression patterns between 2,3,7,8-tetrachlorodibenzo-p-dioxin and a natural aryl hydrocarbon receptor ligand, indirubin. Toxicol Sci. 2004;80:161–169. - PubMed
-
- Ball HJ, Yuasa HJ, Austin CJD, Weiser S, Hunt NH. Indoleamine 2,3-dioxygenase-2: A new enzyme in the kynurenine pathway. Int J Biochem Cell Biol. 2009;41:467–471. - PubMed
-
- Baolin L, Inami Y, Tanaka H, Inagaki N, Iinuma M, Nagai H. Resveratrol inhibits the release of mediators from bone marrow-derived mouse mast cells in vitro. Planta Med. 2004;70:305–309. - PubMed
-
- Beedanagari SR, Bebenek I, Bui P, Hankinson O. Resveratrol inhibits dioxin-induced expression of human CYP1A1 and CYP1B1 by inhibiting recruitment of the aryl hydrocarbon receptor complex and RNA polymerase II to the regulatory regions of the corresponding genes. Toxicol Sci. 2010;116:693–693. - PMC - PubMed
-
- DiNatale BC, Murray IA, Schroeder JC, Flaveny CA, Lahoti TS, Laurenzana EM, Omiecinski CJ, Perdew GH. Kynurenic acid is a potent endogenous aryl hydrocarbon receptor ligand that synergistically induces interleukin-6 in the presence of inflammatory signaling. Toxicol Sci. 2010a;115:89–97. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources