

An examination of the electrostatic interactions between the N-terminal tail of the Brome Mosaic Virus coat protein and encapsidated RNAs
- PMID: 22472420
- PMCID: PMC3360812
- DOI: 10.1016/j.jmb.2012.03.023
An examination of the electrostatic interactions between the N-terminal tail of the Brome Mosaic Virus coat protein and encapsidated RNAs
Abstract
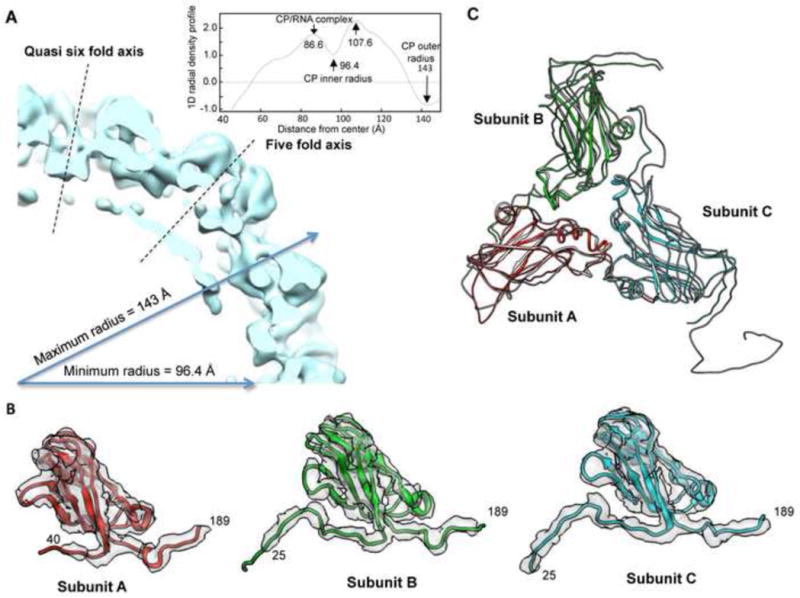
The coat protein of positive-stranded RNA viruses often contains a positively charged tail that extends toward the center of the capsid and interacts with the viral genome. Electrostatic interaction between the tail and the RNA has been postulated as a major force in virus assembly and stabilization. The goal of this work is to examine the correlation between electrostatic interaction and amount of RNA packaged in the tripartite Brome Mosaic Virus (BMV). Nanoindentation experiment using atomic force microscopy showed that the stiffness of BMV virions with different RNAs varied by a range that is 10-fold higher than that would be predicted by electrostatics. BMV mutants with decreased positive charges encapsidated lower amounts of RNA while mutants with increased positive charges packaged additional RNAs up to ∼900 nt. However, the extra RNAs included truncated BMV RNAs, an additional copy of RNA4, potential cellular RNAs, or a combination of the three, indicating that change in the charge of the capsid could result in several different outcomes in RNA encapsidation. In addition, mutant with specific arginines changed to lysines in the capsid also exhibited defects in the specific encapsidation of BMV RNA4. The experimental results indicate that electrostatics is a major component in RNA encapsidation but was unable to account for all of the observed effects on RNA encapsidation. Thermodynamic modeling incorporating the electrostatics was able to predict the approximate length of the RNA to be encapsidated for the majority of mutant virions, but not for a mutant with extreme clustered positive charges. Cryo-electron microscopy of virions that encapsidated an additional copy of RNA4 revealed that, despite the increase in RNA encapsidated, the capsid structure was minimally changed. These results experimentally demonstrated the impact of electrostatics and additional restraints in the encapsidation of BMV RNAs, which could be applicable to other viruses.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures






Similar articles
-
Phosphorylation of the Brome Mosaic Virus Capsid Regulates the Timing of Viral Infection.J Virol. 2016 Aug 12;90(17):7748-60. doi: 10.1128/JVI.00833-16. Print 2016 Sep 1. J Virol. 2016. PMID: 27334588 Free PMC article.
-
The plant host can affect the encapsidation of brome mosaic virus (BMV) RNA: BMV virions are surprisingly heterogeneous.J Mol Biol. 2014 Mar 6;426(5):1061-76. doi: 10.1016/j.jmb.2013.09.007. Epub 2013 Sep 13. J Mol Biol. 2014. PMID: 24036424 Free PMC article.
-
Effects of amino-acid substitutions in the Brome mosaic virus capsid protein on RNA encapsidation.Mol Plant Microbe Interact. 2010 Nov;23(11):1433-47. doi: 10.1094/MPMI-05-10-0118. Mol Plant Microbe Interact. 2010. PMID: 20923351
-
The tripartite virions of the brome mosaic virus have distinct physical properties that affect the timing of the infection process.J Virol. 2014 Jun;88(11):6483-91. doi: 10.1128/JVI.00377-14. Epub 2014 Mar 26. J Virol. 2014. PMID: 24672042 Free PMC article.
-
The coat protein leads the way: an update on basic and applied studies with the Brome mosaic virus coat protein.Mol Plant Pathol. 2011 May;12(4):403-12. doi: 10.1111/j.1364-3703.2010.00678.x. Epub 2010 Nov 25. Mol Plant Pathol. 2011. PMID: 21453435 Free PMC article. Review.
Cited by
-
Phosphorylation of the Brome Mosaic Virus Capsid Regulates the Timing of Viral Infection.J Virol. 2016 Aug 12;90(17):7748-60. doi: 10.1128/JVI.00833-16. Print 2016 Sep 1. J Virol. 2016. PMID: 27334588 Free PMC article.
-
Role of electrostatics in the assembly pathway of a single-stranded RNA virus.J Virol. 2014 Sep;88(18):10472-9. doi: 10.1128/JVI.01044-14. Epub 2014 Jun 25. J Virol. 2014. PMID: 24965458 Free PMC article.
-
Transmissible gastroenteritis coronavirus genome packaging signal is located at the 5' end of the genome and promotes viral RNA incorporation into virions in a replication-independent process.J Virol. 2013 Nov;87(21):11579-90. doi: 10.1128/JVI.01836-13. Epub 2013 Aug 21. J Virol. 2013. PMID: 23966403 Free PMC article.
-
How simple can a model of an empty viral capsid be? Charge distributions in viral capsids.J Biol Phys. 2012 Sep;38(4):657-71. doi: 10.1007/s10867-012-9278-4. Epub 2012 Sep 6. J Biol Phys. 2012. PMID: 24615225 Free PMC article.
-
Virus Assembly Pathways: Straying Away but Not Too Far.Small. 2020 Dec;16(51):e2004475. doi: 10.1002/smll.202004475. Epub 2020 Nov 25. Small. 2020. PMID: 33241653 Free PMC article.
References
-
- Ruigrok RW, Crépin T, Kolakofsky D. Nucleoproteins and nucleocapsids of negative-strand RNA viruses. Curr Opin Microbiol. 2011;14:504–510. - PubMed
-
- Silva AM, Rossmann MG. The Refinement of Southern Bean Mosaic Virus in Reciprocal Space. Acta Crystallogr, Sect B. 1985;41:147–157.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
