

Ovastacin, a cortical granule protease, cleaves ZP2 in the zona pellucida to prevent polyspermy
- PMID: 22472438
- PMCID: PMC3317803
- DOI: 10.1083/jcb.201112094
Ovastacin, a cortical granule protease, cleaves ZP2 in the zona pellucida to prevent polyspermy
Abstract
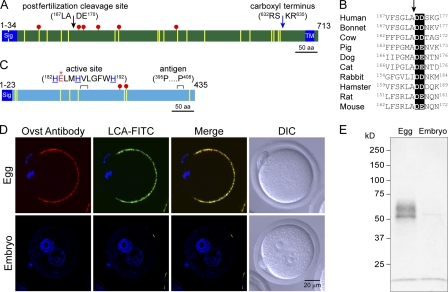
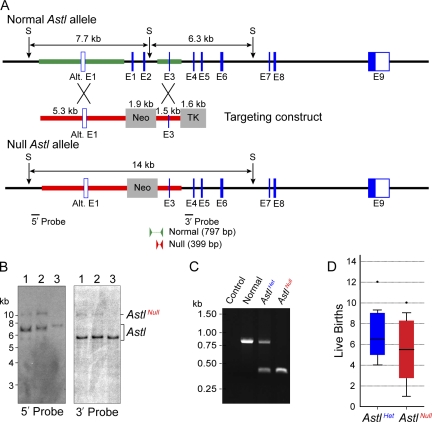
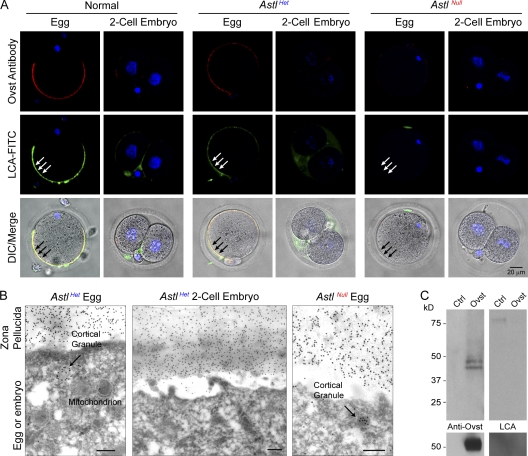
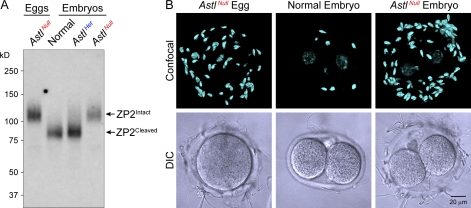
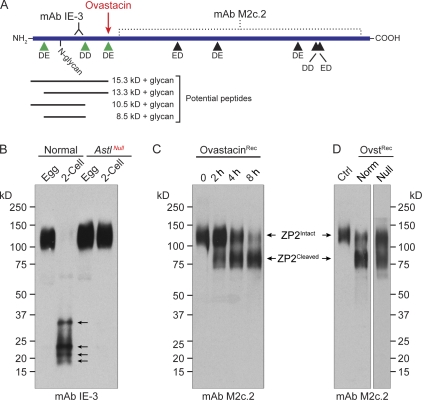
The mouse zona pellucida is composed of three glycoproteins (ZP1, ZP2, and ZP3), of which ZP2 is proteolytically cleaved after gamete fusion to prevent polyspermy. This cleavage is associated with exocytosis of cortical granules that are peripherally located subcellular organelles unique to ovulated eggs. Based on the cleavage site of ZP2, ovastacin was selected as a candidate protease. Encoded by the single-copy Astl gene, ovastacin is an oocyte-specific member of the astacin family of metalloendoproteases. Using specific antiserum, ovastacin was detected in cortical granules before, but not after, fertilization. Recombinant ovastacin cleaved ZP2 in native zonae pellucidae, documenting that ZP2 was a direct substrate of this metalloendoprotease. Female mice lacking ovastacin did not cleave ZP2 after fertilization, and mouse sperm bound as well to Astl-null two-cell embryos as they did to normal eggs. Ovastacin is a pioneer component of mouse cortical granules and plays a definitive role in the postfertilization block to sperm binding that ensures monospermic fertilization and successful development.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
