

Lentiviral vector induced insertional haploinsufficiency of Ebf1 causes murine leukemia
- PMID: 22472950
- PMCID: PMC3369288
- DOI: 10.1038/mt.2012.59
Lentiviral vector induced insertional haploinsufficiency of Ebf1 causes murine leukemia
Abstract
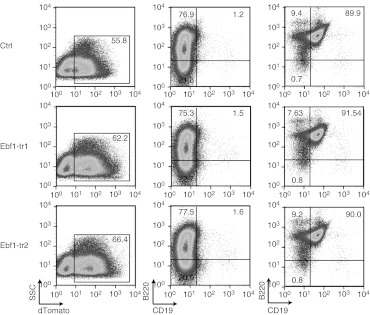
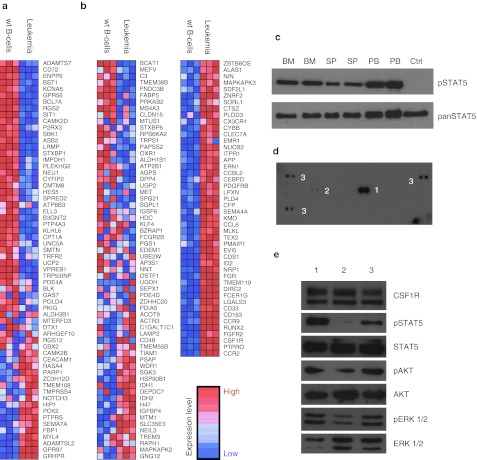
Integrating vectors developed on the basis of various retroviruses have demonstrated therapeutic potential following genetic modification of long-lived hematopoietic stem and progenitor cells. Lentiviral vectors (LV) are assumed to circumvent genotoxic events previously observed with γ-retroviral vectors, due to their integration bias to transcription units in comparison to the γ-retroviral preference for promoter regions and CpG islands. However, recently several studies have revealed the potential for gene activation by LV insertions. Here, we report a murine acute B-lymphoblastic leukemia (B-ALL) triggered by insertional gene inactivation. LV integration occurred into the 8th intron of Ebf1, a major regulator of B-lymphopoiesis. Various aberrant splice variants could be detected that involved splice donor and acceptor sites of the lentiviral construct, inducing downregulation of Ebf1 full-length message. The transcriptome signature was compatible with loss of this major determinant of B-cell differentiation, with partial acquisition of myeloid markers, including Csf1r (macrophage colony-stimulating factor (M-CSF) receptor). This was accompanied by receptor phosphorylation and STAT5 activation, both most likely contributing to leukemic progression. Our results highlight the risk of intragenic vector integration to initiate leukemia by inducing haploinsufficiency of a tumor suppressor gene. We propose to address this risk in future vector design.
Figures
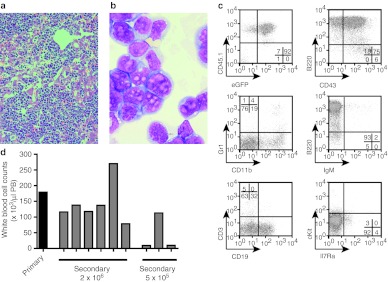
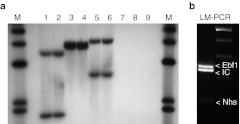
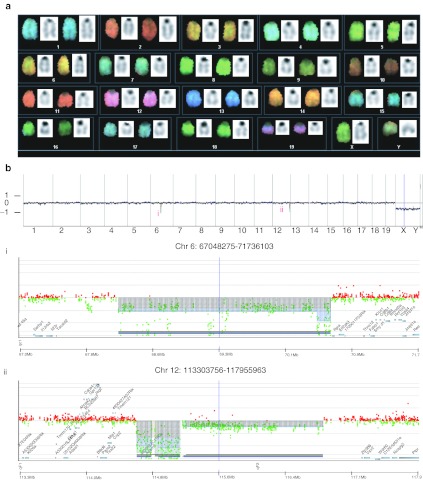
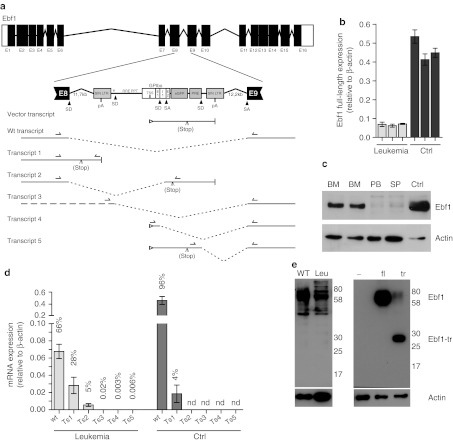
Similar articles
-
Ebf1 or Pax5 haploinsufficiency synergizes with STAT5 activation to initiate acute lymphoblastic leukemia.J Exp Med. 2011 Jun 6;208(6):1135-49. doi: 10.1084/jem.20101947. Epub 2011 May 23. J Exp Med. 2011. PMID: 21606506 Free PMC article.
-
Murine leukemia following irradiation conditioning for transplantation of lentivirally-modified hematopoietic stem cells.Eur J Haematol. 2007 Apr;78(4):303-13. doi: 10.1111/j.1600-0609.2006.00813.x. Eur J Haematol. 2007. PMID: 17378892
-
Preclinical Development of a Hematopoietic Stem and Progenitor Cell Bioengineered Factor VIII Lentiviral Vector Gene Therapy for Hemophilia A.Hum Gene Ther. 2018 Oct;29(10):1183-1201. doi: 10.1089/hum.2018.137. Hum Gene Ther. 2018. PMID: 30160169 Free PMC article.
-
Understanding lentiviral vector chromatin targeting: working to reduce insertional mutagenic potential for gene therapy.Gene Ther. 2013 Jun;20(6):581-8. doi: 10.1038/gt.2012.88. Epub 2012 Nov 22. Gene Ther. 2013. PMID: 23171920 Review.
-
[Progress in improvement of lentiviral vectors' transcriptional read-through].Sheng Wu Gong Cheng Xue Bao. 2011 Nov;27(11):1541-8. Sheng Wu Gong Cheng Xue Bao. 2011. PMID: 22393708 Review. Chinese.
Cited by
-
Long-term tracking of segmental bone healing mediated by genetically engineered adipose-derived stem cells: focuses on bone remodeling and potential side effects.Tissue Eng Part A. 2014 May;20(9-10):1392-402. doi: 10.1089/ten.TEA.2013.0314. Tissue Eng Part A. 2014. PMID: 24367947 Free PMC article.
-
Alpharetroviral vectors: from a cancer-causing agent to a useful tool for human gene therapy.Viruses. 2014 Dec 5;6(12):4811-38. doi: 10.3390/v6124811. Viruses. 2014. PMID: 25490763 Free PMC article. Review.
-
Steerable Induction of the Thymosin β4/MRTF-A Pathway via AAV-Based Overexpression Induces Therapeutic Neovascularization.Hum Gene Ther. 2017 Oct 3;29(12):1407-15. doi: 10.1089/hum.2017.013. Online ahead of print. Hum Gene Ther. 2017. PMID: 28726522 Free PMC article.
-
Cancer gene discovery: exploiting insertional mutagenesis.Mol Cancer Res. 2013 Oct;11(10):1141-58. doi: 10.1158/1541-7786.MCR-13-0244. Epub 2013 Aug 8. Mol Cancer Res. 2013. PMID: 23928056 Free PMC article. Review.
-
Enhanced Transduction of Human Hematopoietic Stem Cells by AAV6 Vectors: Implications in Gene Therapy and Genome Editing.Mol Ther Nucleic Acids. 2020 Jun 5;20:451-458. doi: 10.1016/j.omtn.2020.03.009. Epub 2020 Mar 29. Mol Ther Nucleic Acids. 2020. PMID: 32276210 Free PMC article.
References
-
- Qasim W, Gaspar HB., and, Thrasher AJ. Progress and prospects: gene therapy for inherited immunodeficiencies. Gene Ther. 2009;16:1285–1291. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
