

Essential role of EBF1 in the generation and function of distinct mature B cell types
- PMID: 22473956
- PMCID: PMC3328360
- DOI: 10.1084/jem.20112422
Essential role of EBF1 in the generation and function of distinct mature B cell types
Abstract
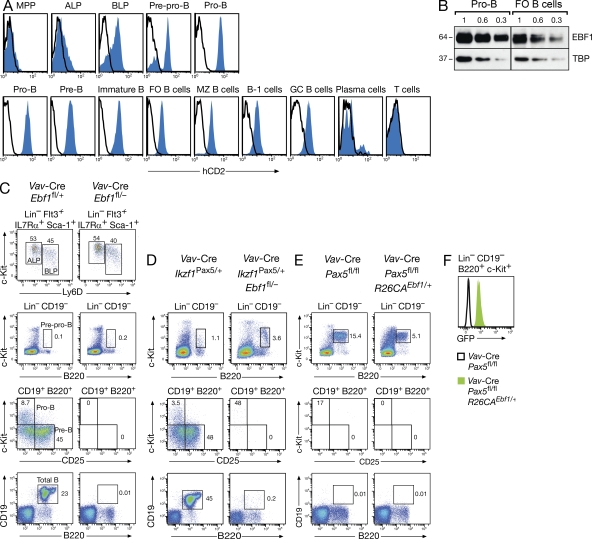
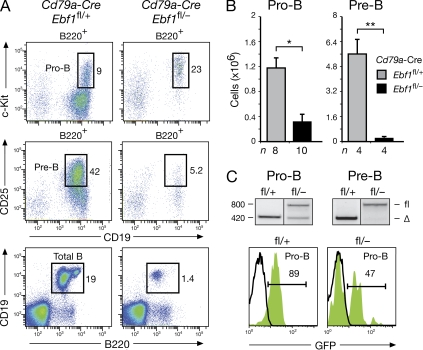
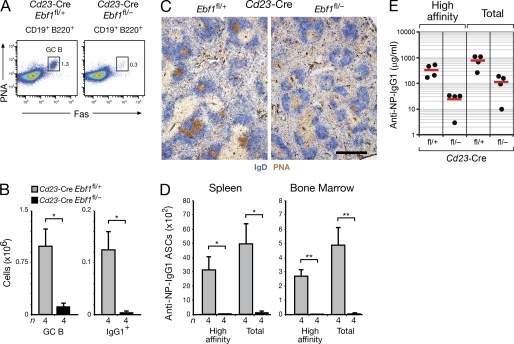
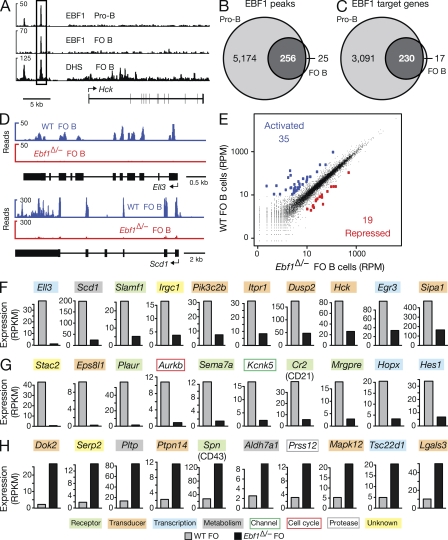
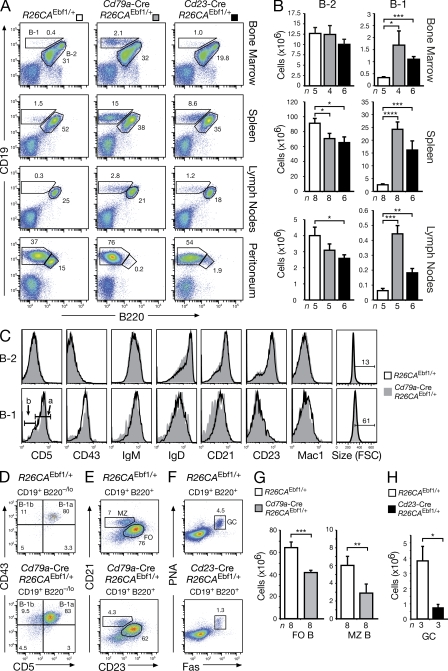
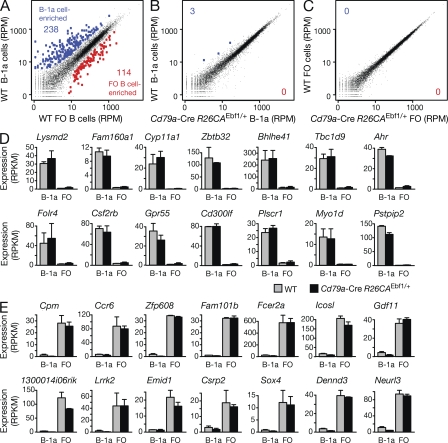
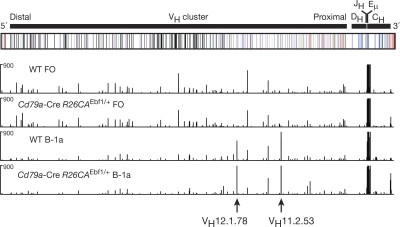
The transcription factor EBF1 is essential for lineage specification in early B cell development. In this study, we demonstrate by conditional mutagenesis that EBF1 is required for B cell commitment, pro-B cell development, and subsequent transition to the pre-B cell stage. Later in B cell development, EBF1 was essential for the generation and maintenance of several mature B cell types. Marginal zone and B-1 B cells were lost, whereas follicular (FO) and germinal center (GC) B cells were reduced in the absence of EBF1. Activation of the B cell receptor resulted in impaired intracellular signaling, proliferation and survival of EBF1-deficient FO B cells. Immune responses were severely reduced upon Ebf1 inactivation, as GCs were formed but not maintained. ChIP- and RNA-sequencing of FO B cells identified EBF1-activated genes that encode receptors, signal transducers, and transcriptional regulators implicated in B cell signaling. Notably, ectopic expression of EBF1 efficiently induced the development of B-1 cells at the expense of conventional B cells. These gain- and loss-of-function analyses uncovered novel important functions of EBF1 in controlling B cell immunity.
Figures



Similar articles
-
The regulatory network of B-cell differentiation: a focused view of early B-cell factor 1 function.Immunol Rev. 2014 Sep;261(1):102-15. doi: 10.1111/imr.12206. Immunol Rev. 2014. PMID: 25123279 Free PMC article. Review.
-
Transcription factor Ebf1 regulates differentiation stage-specific signaling, proliferation, and survival of B cells.Genes Dev. 2012 Apr 1;26(7):668-82. doi: 10.1101/gad.187328.112. Epub 2012 Mar 19. Genes Dev. 2012. PMID: 22431510 Free PMC article.
-
Combined heterozygous loss of Ebf1 and Pax5 allows for T-lineage conversion of B cell progenitors.J Exp Med. 2015 Jun 29;212(7):1109-23. doi: 10.1084/jem.20132100. Epub 2015 Jun 8. J Exp Med. 2015. PMID: 26056231 Free PMC article.
-
Distinct promoters mediate the regulation of Ebf1 gene expression by interleukin-7 and Pax5.Mol Cell Biol. 2007 Jan;27(2):579-94. doi: 10.1128/MCB.01192-06. Epub 2006 Nov 13. Mol Cell Biol. 2007. PMID: 17101802 Free PMC article.
-
Essential functions of Pax-5 (BSAP) in pro-B cell development.Immunobiology. 1997 Dec;198(1-3):227-35. doi: 10.1016/S0171-2985(97)80043-5. Immunobiology. 1997. PMID: 9442394 Review.
Cited by
-
A capture-sequencing strategy identifies IRF8, EBF1, and APRIL as novel IGH fusion partners in B-cell lymphoma.Blood. 2013 Aug 1;122(5):726-33. doi: 10.1182/blood-2013-04-495804. Epub 2013 Jun 17. Blood. 2013. PMID: 23775715 Free PMC article.
-
Impaired Cellular Immunity in the Murine Neural Crest Conditional Deletion of Endothelin Receptor-B Model of Hirschsprung's Disease.PLoS One. 2015 Jun 10;10(6):e0128822. doi: 10.1371/journal.pone.0128822. eCollection 2015. PLoS One. 2015. PMID: 26061883 Free PMC article.
-
The emerging role of Epstein-Barr virus encoded microRNAs in nasopharyngeal carcinoma.J Cancer. 2018 Jul 30;9(16):2852-2864. doi: 10.7150/jca.25460. eCollection 2018. J Cancer. 2018. PMID: 30123354 Free PMC article. Review.
-
IL-7R signaling activates widespread VH and DH gene usage to drive antibody diversity in bone marrow B cells.Cell Rep. 2021 Jul 13;36(2):109349. doi: 10.1016/j.celrep.2021.109349. Cell Rep. 2021. PMID: 34260907 Free PMC article.
-
The regulatory network of B-cell differentiation: a focused view of early B-cell factor 1 function.Immunol Rev. 2014 Sep;261(1):102-15. doi: 10.1111/imr.12206. Immunol Rev. 2014. PMID: 25123279 Free PMC article. Review.
References
-
- Ahearn J.M., Fischer M.B., Croix D., Goerg S., Ma M., Xia J., Zhou X., Howard R.G., Rothstein T.L., Carroll M.C. 1996. Disruption of the Cr2 locus results in a reduction in B-1a cells and in an impaired B cell response to T-dependent antigen. Immunity. 4:251–262 10.1016/S1074-7613(00)80433-1 - DOI - PubMed
-
- Bain G., Maandag E.C.R., Izon D.J., Amsen D., Kruisbeek A.M., Weintraub B.C., Krop I., Schlissel M.S., Feeney A.J., van Roon M., et al. 1994. E2A proteins are required for proper B cell development and initiation of immunoglobulin gene rearrangements. Cell. 79:885–892 10.1016/0092-8674(94)90077-9 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
