

Bile acids down-regulate caveolin-1 in esophageal epithelial cells through sterol responsive element-binding protein
- PMID: 22474125
- PMCID: PMC5417097
- DOI: 10.1210/me.2011-1140
Bile acids down-regulate caveolin-1 in esophageal epithelial cells through sterol responsive element-binding protein
Abstract
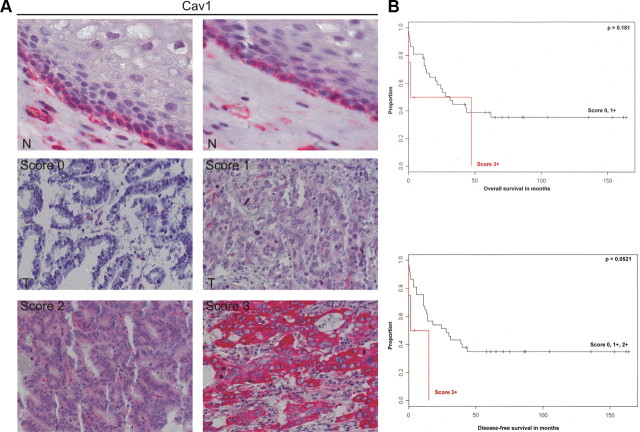
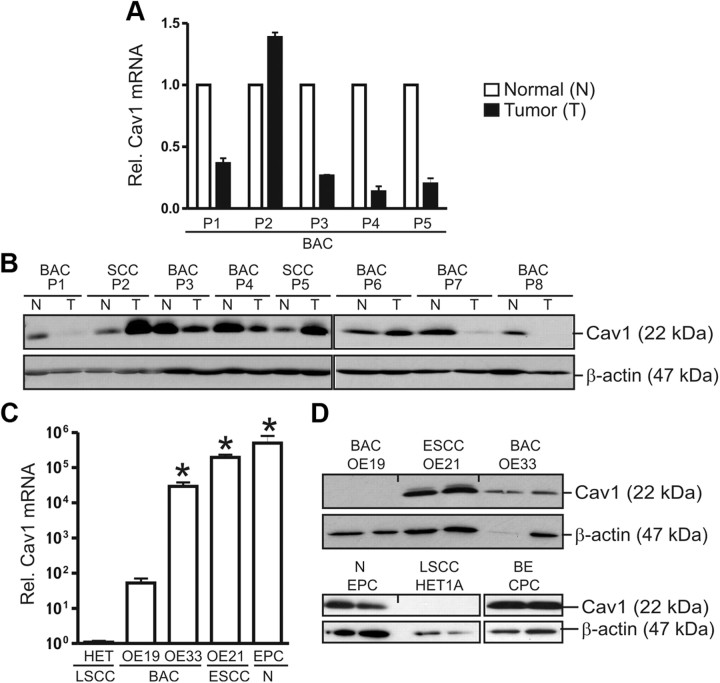
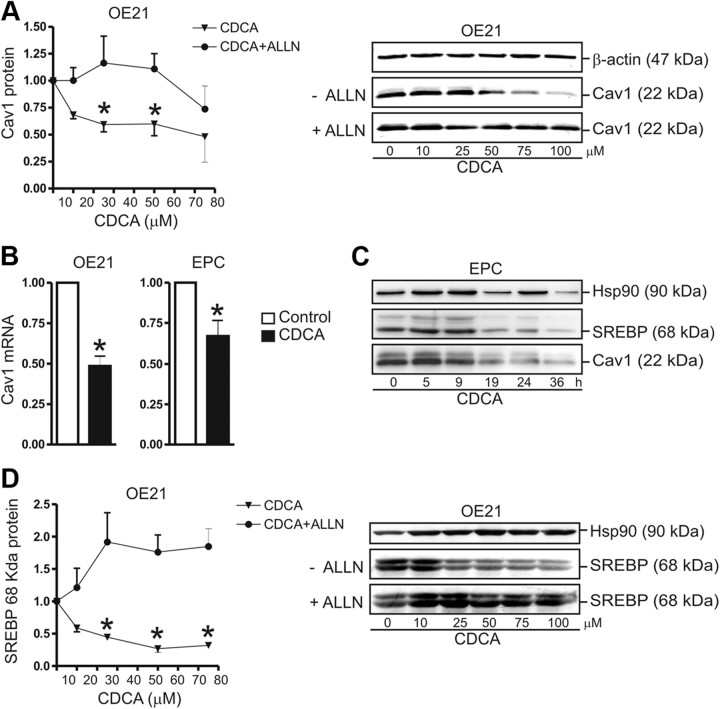
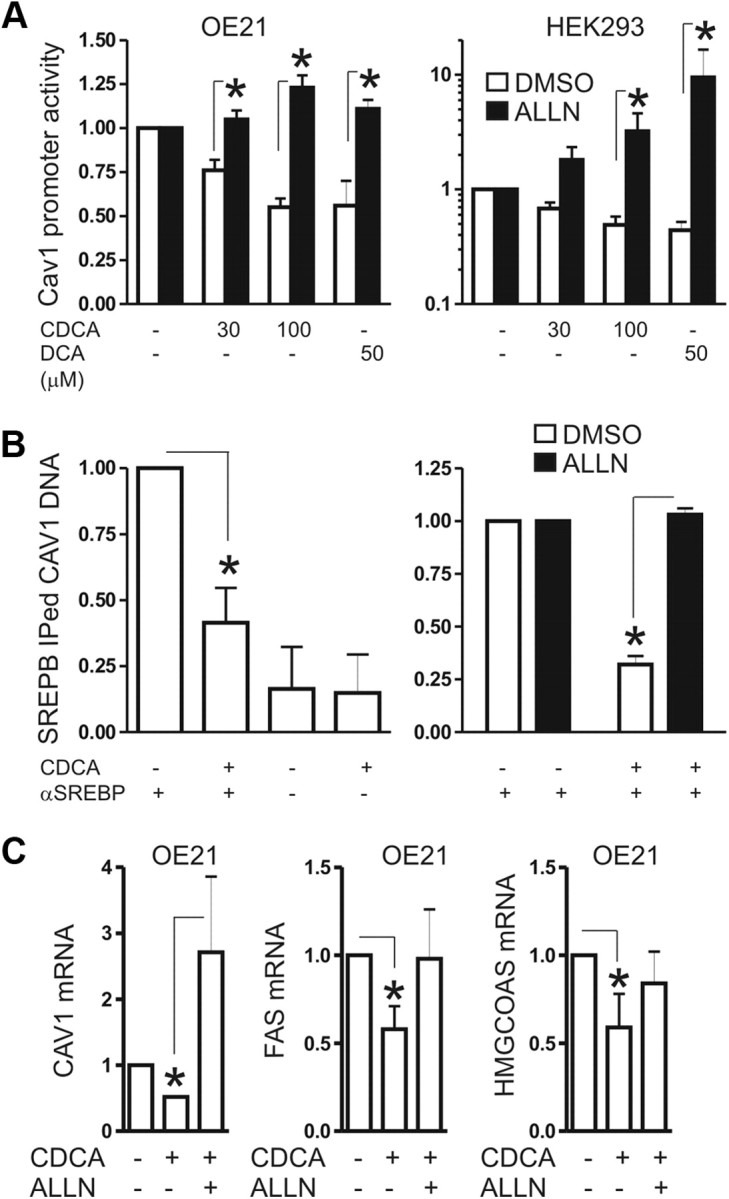
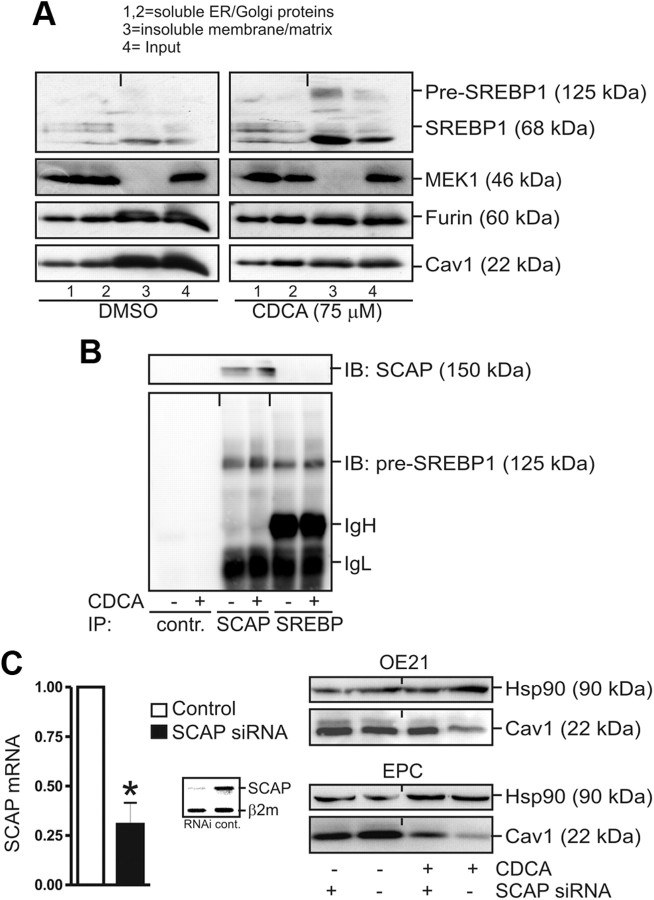
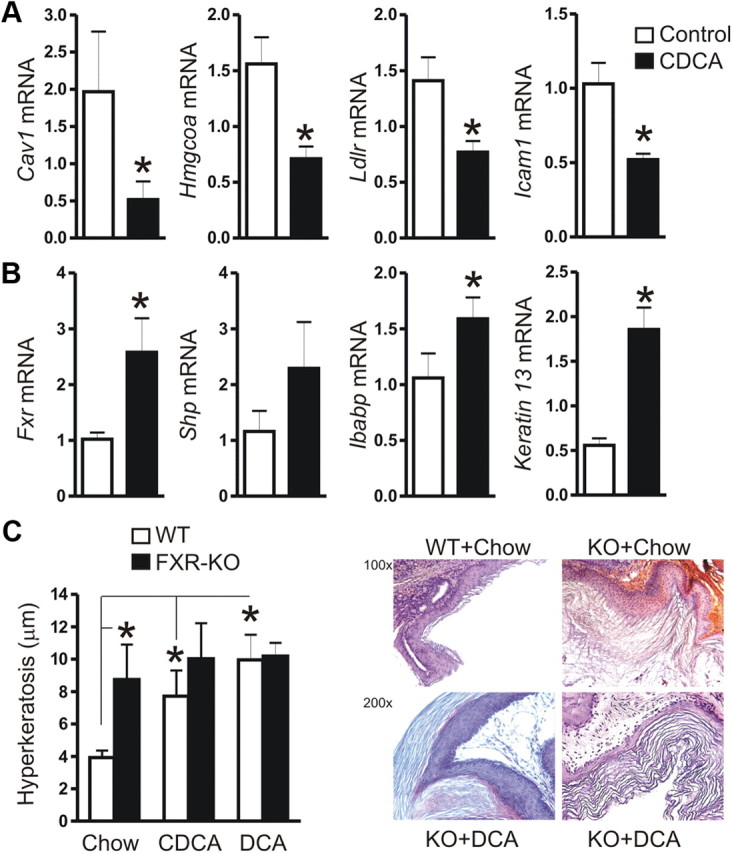
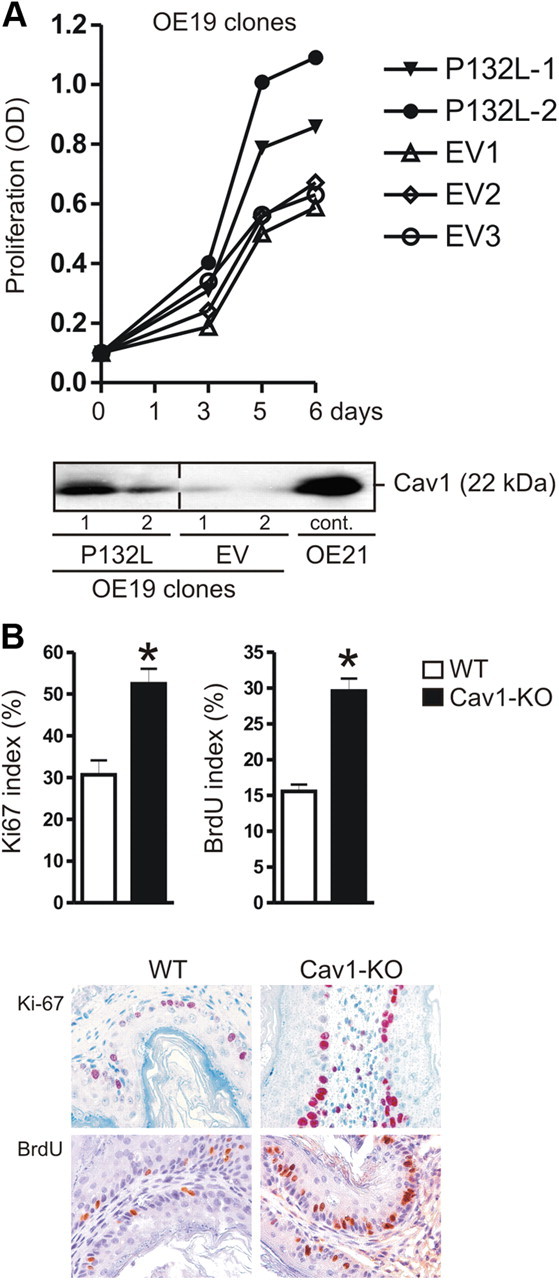
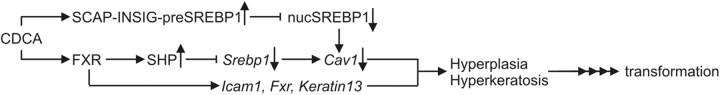
Bile acids are synthesized from cholesterol and are major risk factors for Barrett adenocarcinoma (BAC) of the esophagus. Caveolin-1 (Cav1), a scaffold protein of membrane caveolae, is transcriptionally regulated by cholesterol via sterol-responsive element-binding protein-1 (SREBP1). Cav1 protects squamous epithelia by controlling cell growth and stabilizing cell junctions and matrix adhesion. Cav1 is frequently down-regulated in human cancers; however, the molecular mechanisms that lead to this event are unknown. We show that the basal layer of the nonneoplastic human esophageal squamous epithelium expressed Cav1 mainly at intercellular junctions. In contrast, Cav1 was lost in 95% of tissue specimens from BAC patients (n = 100). A strong cytoplasmic expression of Cav1 correlated with poor survival in a small subgroup (n = 5) of BAC patients, and stable expression of an oncogenic Cav1 variant (Cav1-P132L) in the human BAC cell line OE19 promoted proliferation. Cav1 was also detectable in immortalized human squamous epithelial, Barrett esophagus (CPC), and squamous cell carcinoma cells (OE21), but was low in BAC cell lines (OE19, OE33). Mechanistically, bile acids down-regulated Cav1 expression by inhibition of the proteolytic cleavage of 125-kDa pre-SREBP1 from the endoplasmic reticulum/Golgi apparatus and nuclear translocation of active 68-kDa SREBP1. This block in SREBP1's posttranslational processing impaired transcriptional activation of SREBP1 response elements in the proximal human Cav1 promoter. Cav1 was also down-regulated in esophagi from C57BL/6 mice on a diet enriched with 1% (wt/wt) chenodeoxycholic acid. Mice deficient for Cav1 or the nuclear bile acid receptor farnesoid X receptor showed hyperplasia and hyperkeratosis of the basal cell layer of esophageal epithelia, respectively. These data indicate that bile acid-mediated down-regulation of Cav1 marks early changes in the squamous epithelium, which may contribute to onset of Barrett esophagus metaplasia and progression to BAC.
Figures
References
-
- Shaheen NJ. 2005. Advances in Barrett's esophagus and esophageal adenocarcinoma. Gastroenterology 128:1554–1566 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
