

FGF21 is increased by inflammatory stimuli and protects leptin-deficient ob/ob mice from the toxicity of sepsis
- PMID: 22474187
- PMCID: PMC3359613
- DOI: 10.1210/en.2011-1496
FGF21 is increased by inflammatory stimuli and protects leptin-deficient ob/ob mice from the toxicity of sepsis
Abstract
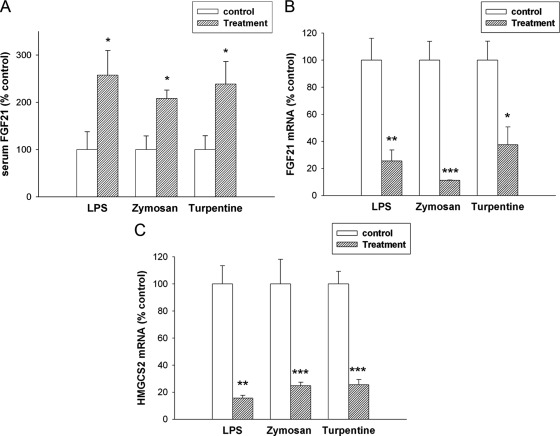
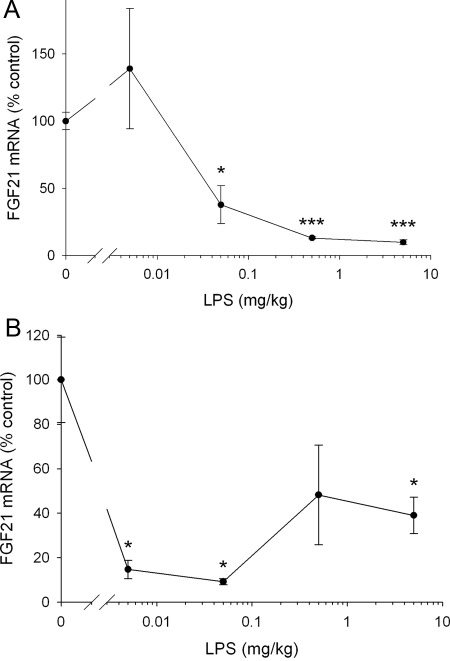
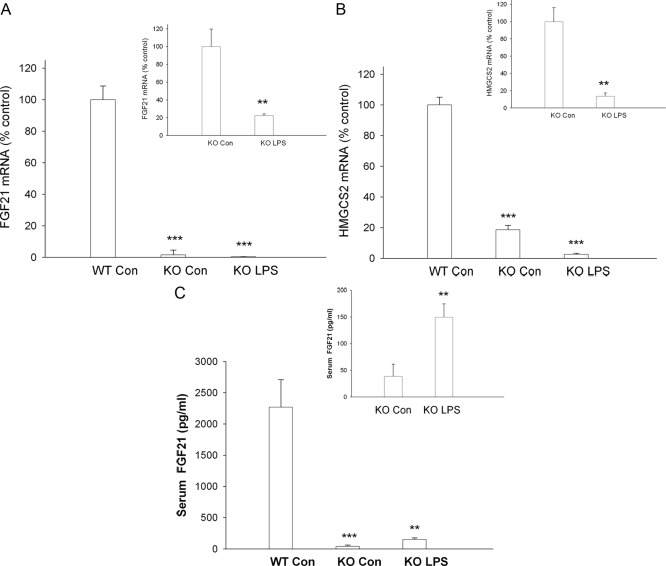
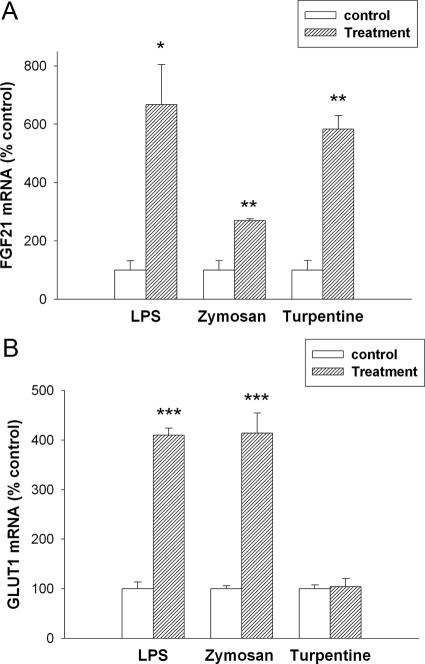
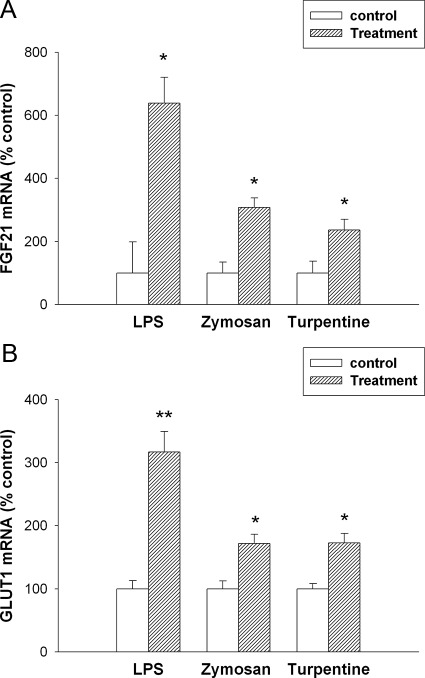
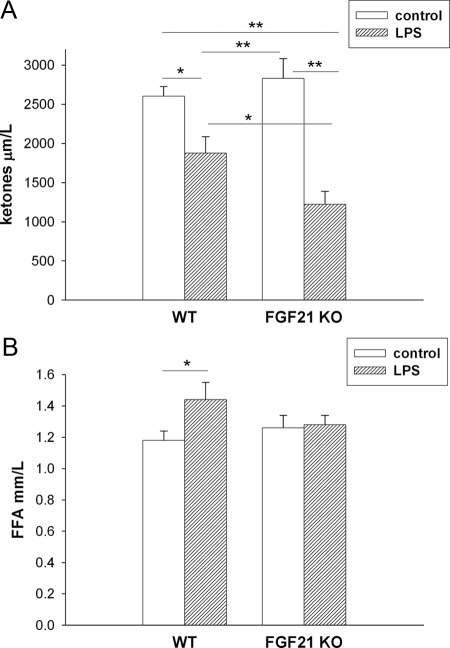
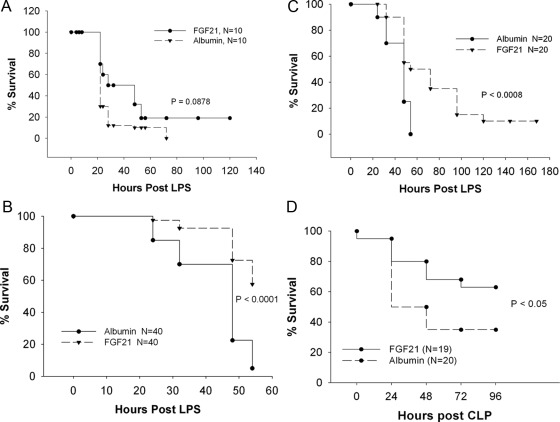
The acute phase response (APR) produces marked alterations in lipid and carbohydrate metabolism including decreasing plasma ketone levels. Fibroblast growth factor 21 (FGF21) is a recently discovered hormone that regulates lipid and glucose metabolism and stimulates ketogenesis. Here we demonstrate that lipopolysaccharide (LPS), zymosan, and turpentine, which induce the APR, increase serum FGF21 levels 2-fold. Although LPS, zymosan, and turpentine decrease the hepatic expression of FGF21, they increase FGF21 expression in adipose tissue and muscle, suggesting that extrahepatic tissues account for the increase in serum FGF21. After LPS administration, the characteristic decrease in plasma ketone levels is accentuated in FGF21-/- mice, but this is not due to differences in expression of carnitine palmitoyltransferase 1α or hydroxymethyglutaryl-CoA synthase 2 in liver, because LPS induces similar decreases in the expression of these genes in FGF21-/- and control mice. However, in FGF21-/- mice, the ability of LPS to increase plasma free fatty acid levels is blunted. This failure to increase plasma free fatty acid could contribute to the accentuated decrease in plasma ketone levels because the transport of fatty acids from adipose tissue to liver provides the substrate for ketogenesis. Treatment with exogenous FGF21 reduced the number of animals that die and the rapidity of death after LPS administration in leptin-deficient ob/ob mice and to a lesser extent in control mice. FGF21 also protected from the toxic effects of cecal ligation and puncture-induced sepsis. Thus, FGF21 is a positive APR protein that protects animals from the toxic effects of LPS and sepsis.
Figures
References
-
- Baumann H, Gauldie J. 1994. The acute phase response. Immunol Today 15:74–80 - PubMed
-
- Gabay C, Kushner I. 1999. Acute-phase proteins and other systemic responses to inflammation. N Engl J Med 340:448–454 - PubMed
-
- Khovidhunkit W, Kim MS, Memon RA, Shigenaga JK, Moser AH, Feingold KR, Grunfeld C. 2004. Effects of infection and inflammation on lipid and lipoprotein metabolism: mechanisms and consequences to the host. J Lipid Res 45:1169–1196 - PubMed
-
- Spitzer JJ. 1995. Bacterial endotoxin effects on carbohydrate utilization and transport. Biochem Soc Trans 23:998–1002 - PubMed
-
- Wolfe RR. 1997. Substrate utilization/insulin resistance in sepsis/trauma. Baillieres Clin Endocrinol Metab 11:645–657 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
