

Hedgehog secretion and signal transduction in vertebrates
- PMID: 22474285
- PMCID: PMC3365689
- DOI: 10.1074/jbc.R112.356006
Hedgehog secretion and signal transduction in vertebrates
Abstract
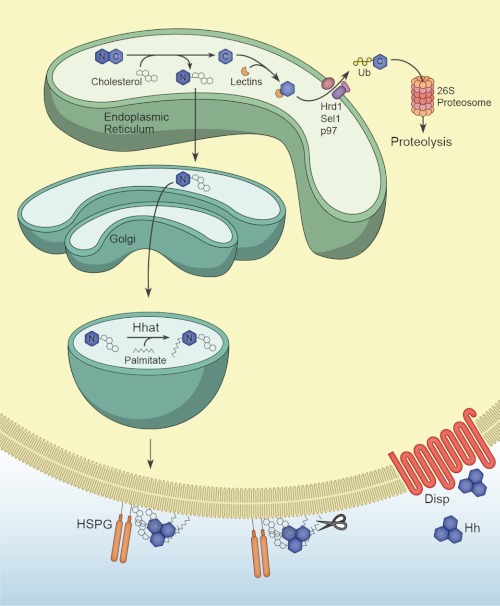
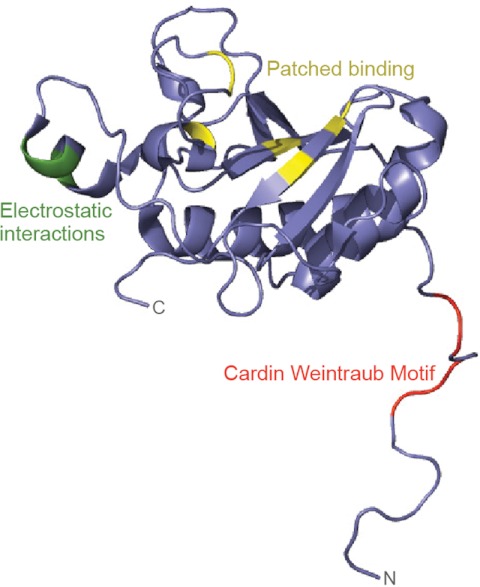
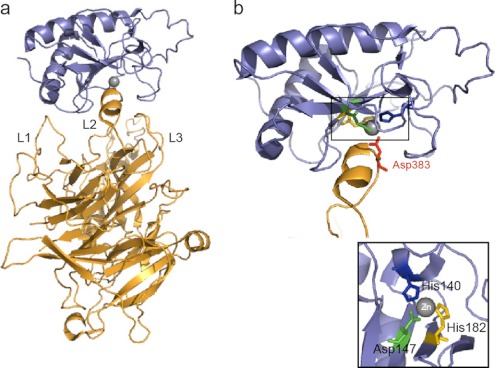
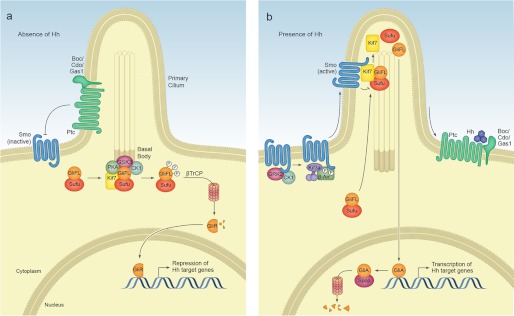
Signaling by the Hedgehog (Hh) family of secreted proteins is essential for proper embryonic patterning and development. Dysregulation of Hh signaling is associated with a variety of human diseases ranging from developmental disorders such as holoprosencephaly to certain forms of cancer, including medulloblastoma and basal cell carcinoma. Genetic studies in flies and mice have shaped our understanding of Hh signaling and revealed that nearly all core components of the pathway are highly conserved. Although many aspects of the Drosophila Hh pathway are conserved in vertebrates, mechanistic differences between the two species have begun to emerge. Perhaps the most striking divergence in vertebrate Hh signaling is its dependence on the primary cilium, a vestigial organelle that is largely absent in flies. This minireview will provide an overview of Hh signaling and present recent insights into vertebrate Hh secretion, receptor binding, and signal transduction.
Figures
References
-
- Ingham P. W., McMahon A. P. (2001) Hedgehog signaling in animal development: paradigms and principles. Genes Dev. 15, 3059–3087 - PubMed
-
- Ingham P. W., Nakano Y., Seger C. (2011) Mechanisms and functions of Hedgehog signaling across the metazoa. Nat. Rev. Genet 12, 393–406 - PubMed
-
- Pedersen L. B., Rosenbaum J. L. (2008) Intraflagellar transport (IFT) role in ciliary assembly, resorption, and signaling. Curr. Top. Dev. Biol. 85, 23–61 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
