

Viral-induced encephalitis initiates distinct and functional CD103+ CD11b+ brain dendritic cell populations within the olfactory bulb
- PMID: 22474352
- PMCID: PMC3340999
- DOI: 10.1073/pnas.1203941109
Viral-induced encephalitis initiates distinct and functional CD103+ CD11b+ brain dendritic cell populations within the olfactory bulb
Abstract
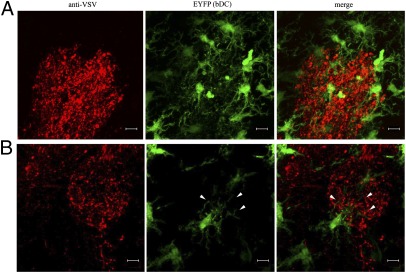
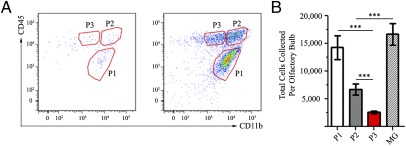
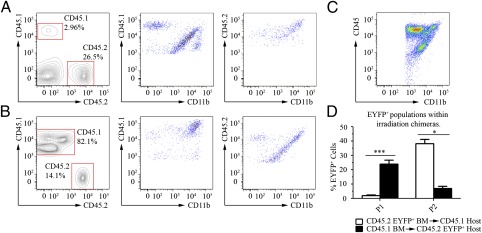
Dendritic cells (DC) are antigen-presenting cells found in both lymphoid and nonlymphoid organs, including the brain (bDC) of Cd11c/eyfp transgenic C57BL/6 mice. Using an intranasal vesicular stomatitis virus infection, we demonstrated that EYFP(+) cells amass in areas associated with viral antigens, take on an activated morphology, and project their processes into infected neuronal tissue within the olfactory bulb. These bDC separated into three EYFP(+) CD45(+) CD11b(+) populations, all but one being able to functionally promote both T lymphocyte proliferation and T(H)1 cytokine production. One population was shown to emanate from the brain and a second population was peripherally derived. The third population was of indeterminate origin, being both radiosensitive and not replenished by donor bone marrow. Finally, each EYFP(+) population contained CD11b(+) CD103(+) subpopulations and could be distinguished in terms of CD115, Gr-1, and Ly-6C expression, highlighting mucosal and monocyte-derived DC lineages.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Steinman RM, Idoyaga J. Features of the dendritic cell lineage. Immunol Rev. 2010;234:5–17. - PubMed
-
- Steinman RM, et al. Dendritic cell function in vivo during the steady state: A role in peripheral tolerance. Ann N Y Acad Sci. 2003;987:15–25. - PubMed
-
- Steinman RM, Hawiger D, Nussenzweig MC. Tolerogenic dendritic cells. Annu Rev Immunol. 2003;21:685–711. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
