

Structural and mechanistic analysis of the membrane-embedded glycosyltransferase WaaA required for lipopolysaccharide synthesis
- PMID: 22474366
- PMCID: PMC3341020
- DOI: 10.1073/pnas.1119894109
Structural and mechanistic analysis of the membrane-embedded glycosyltransferase WaaA required for lipopolysaccharide synthesis
Abstract
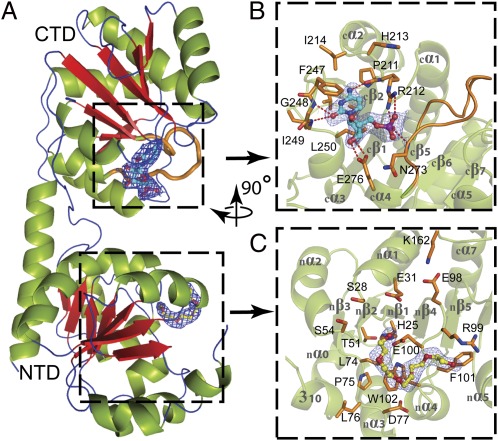
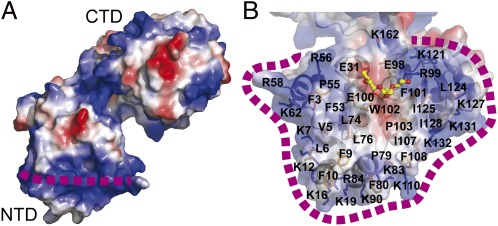
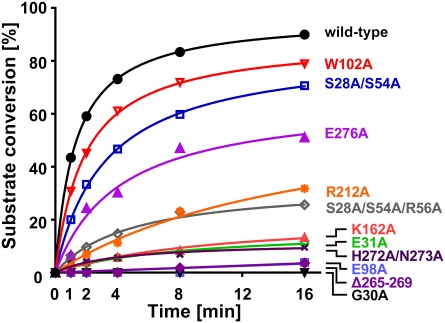
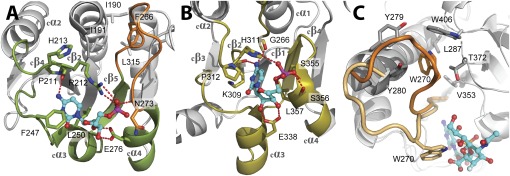
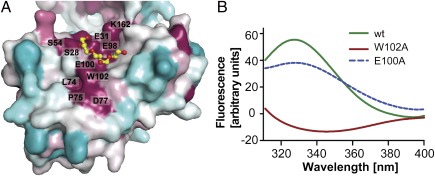
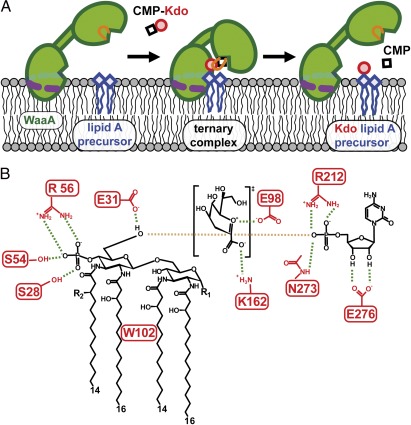
WaaA is a key enzyme in the biosynthesis of LPS, a critical component of the outer envelope of Gram-negative bacteria. Embedded in the cytoplasmic face of the inner membrane, WaaA catalyzes the transfer of 3-deoxy-d-manno-oct-2-ulosonic acid (Kdo) to the lipid A precursor of LPS. Here we present crystal structures of the free and CMP-bound forms of WaaA from Aquifex aeolicus, an ancient Gram-negative hyperthermophile. These structures reveal details of the CMP-binding site and implicate a unique sequence motif (GGS/TX(5)GXNXLE) in Kdo binding. In addition, a cluster of highly conserved amino acid residues was identified which represents the potential membrane-attachment and acceptor-substrate binding site of WaaA. A series of site-directed mutagenesis experiments revealed critical roles for glycine 30 and glutamate 31 in Kdo transfer. Our results provide the structural basis of a critical reaction in LPS biosynthesis and allowed the development of a detailed model of the catalytic mechanism of WaaA.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Wiese A, Brandenburg K, Ulmer AJ, Seydel U, Müller-Loennies S. The dual role of lipopolysaccharide as effector and target molecule. Biol Chem. 1999;380:767–784. - PubMed
-
- Holst O. The structures of core regions from enterobacterial lipopolysaccharides - an update. FEMS Microbiol Lett. 2007;271:3–11. - PubMed
-
- Belunis CJ, Clementz T, Carty SM, Raetz CR. Inhibition of lipopolysaccharide biosynthesis and cell growth following inactivation of the kdtA gene in Escherichia coli. J Biol Chem. 1995;270:27646–27652. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
