

NSP4, an elastase-related protease in human neutrophils with arginine specificity
- PMID: 22474388
- PMCID: PMC3341072
- DOI: 10.1073/pnas.1200470109
NSP4, an elastase-related protease in human neutrophils with arginine specificity
Abstract
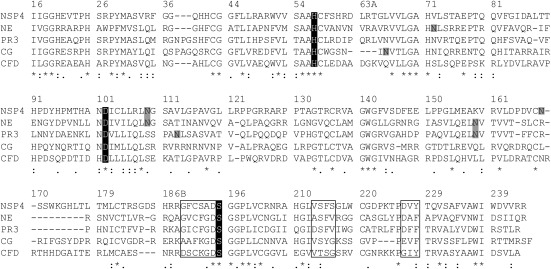
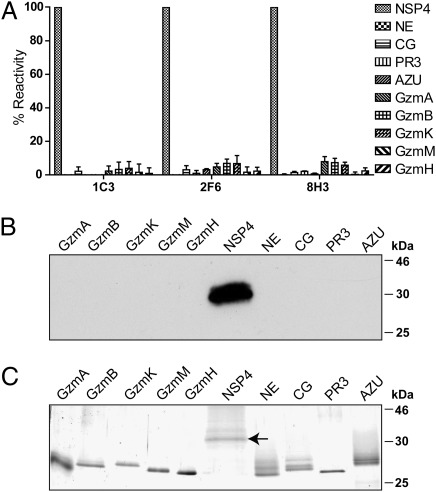
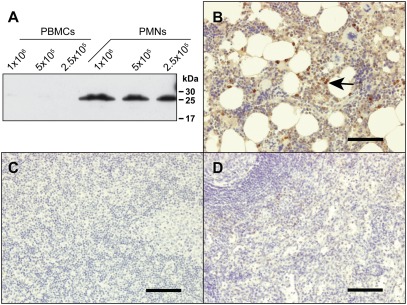
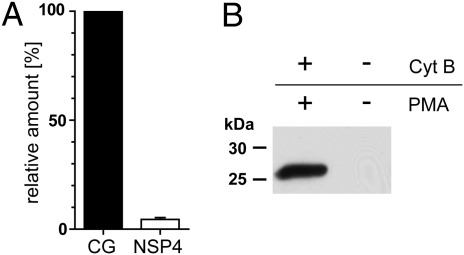
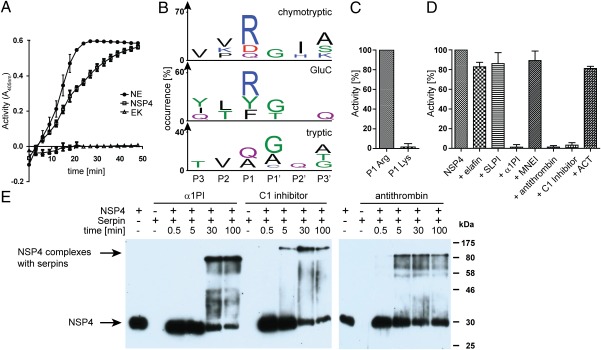
Neutrophil serine proteases (NSPs) in cytoplasmic granules of neutrophils are regarded as important antimicrobial defense weapons after engulfment and exposure of pathogens to the content of primary granules. Despite intensive studies on neutrophils during the last three decades, only three active serine proteases, neutrophil elastase (NE), cathepsin G (CG), and proteinase 3 (PR3) have been identified in these short-lived cells. Here, we report on the identification of a fourth serine protease (NSP4) with 39% identity to NE and PR3, but arginine specificity, yet sharing features like propeptide processing by dipeptidyl peptidase I, storage, and release as an active enzyme with the three active proteases. We established monoclonal antibodies against NSP4, excluded cross-reactivity to human granzymes, NE, CG, PR3, and azurocidin, and screened for NSP4 protein expression in various human tissues and blood leukocyte populations. Only granulocyte precursors and neutrophil populations from peripheral blood were positive. The content of NSP4 in neutrophil lysates, however, was about 20-fold lower compared with CG. Upon neutrophil activation, NSP4 was released into the supernatant. Profiling its specificity with peptide libraries from Escherichia coli revealed a preference for arginine in P1; it cleaved Tyr-Arg-Phe-Arg-AMC and Ala-Pro-Nva-thiobenzyl esters. NSP4 was inhibited by α(1)-proteinase inhibitor (α(1)-antitrypsin), C1 inhibitor, and most efficiently by antithrombin-heparin, but not by elafin, secretory leukocyte protease inhibitor, α(1)-antichymotrypsin, and monocyte-neutrophil elastase inhibitor. Functional specialization and preferred natural substrates of NSP4 remain to be determined to understand the biological interplay of all four NSPs during neutrophil responses.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
