

Adaptive immune response to model antigens is impaired in murine leukocyte-adhesion deficiency-1 revealing elevated activation thresholds in vivo
- PMID: 22474478
- PMCID: PMC3303708
- DOI: 10.1155/2012/450738
Adaptive immune response to model antigens is impaired in murine leukocyte-adhesion deficiency-1 revealing elevated activation thresholds in vivo
Abstract
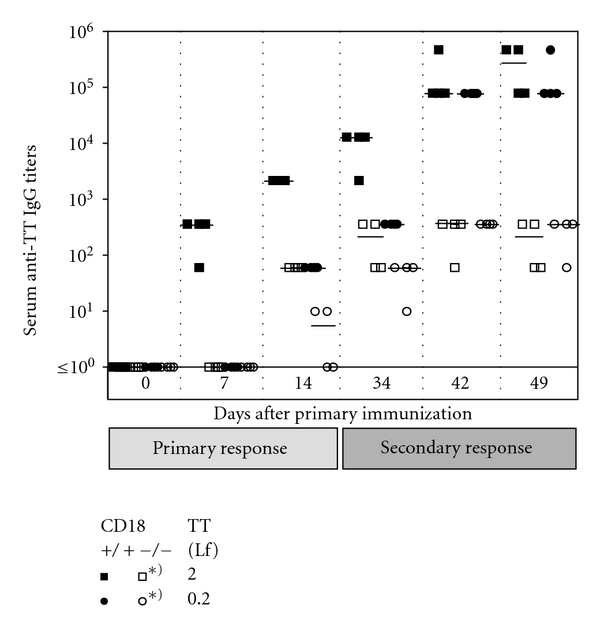
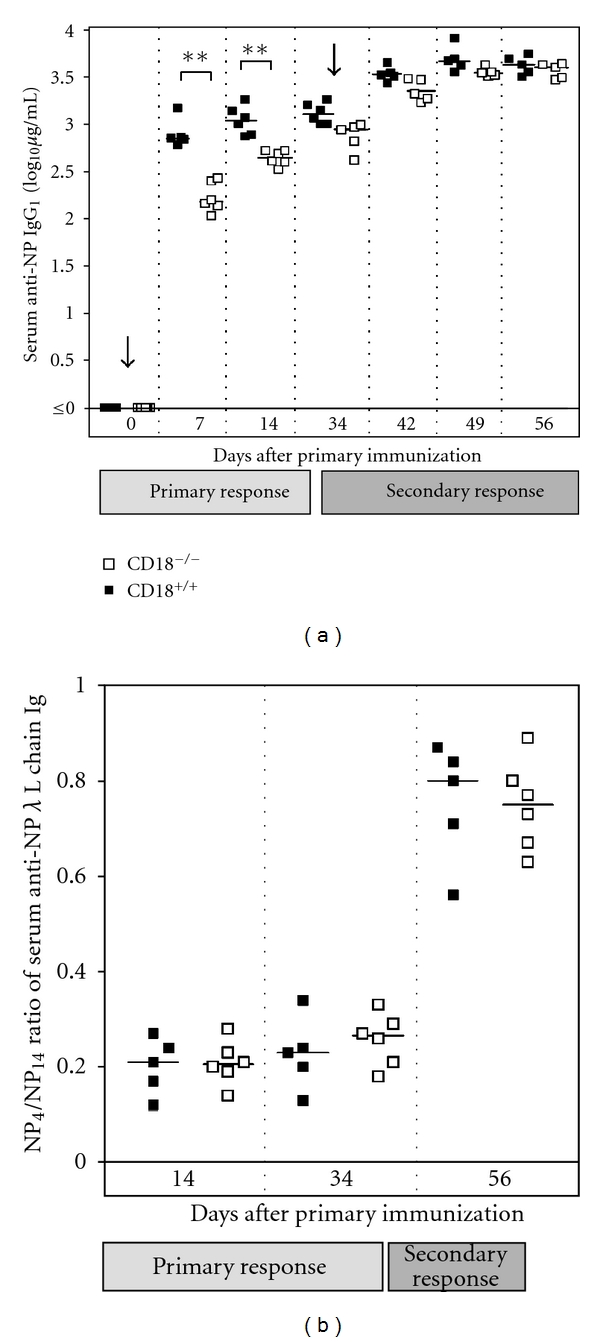
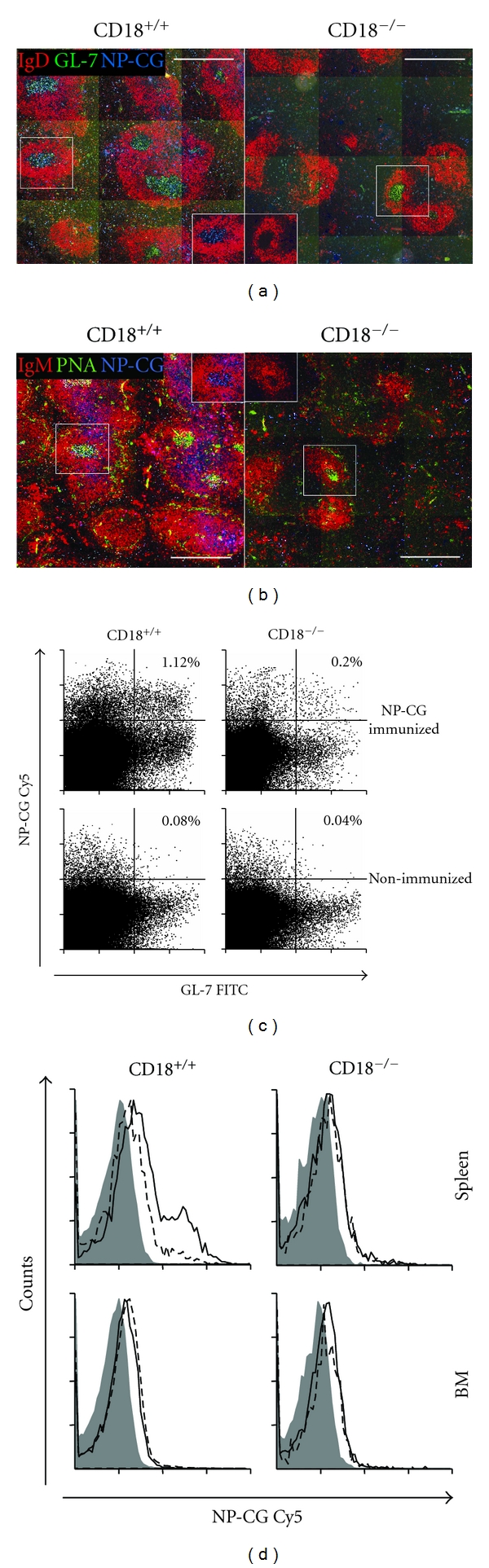
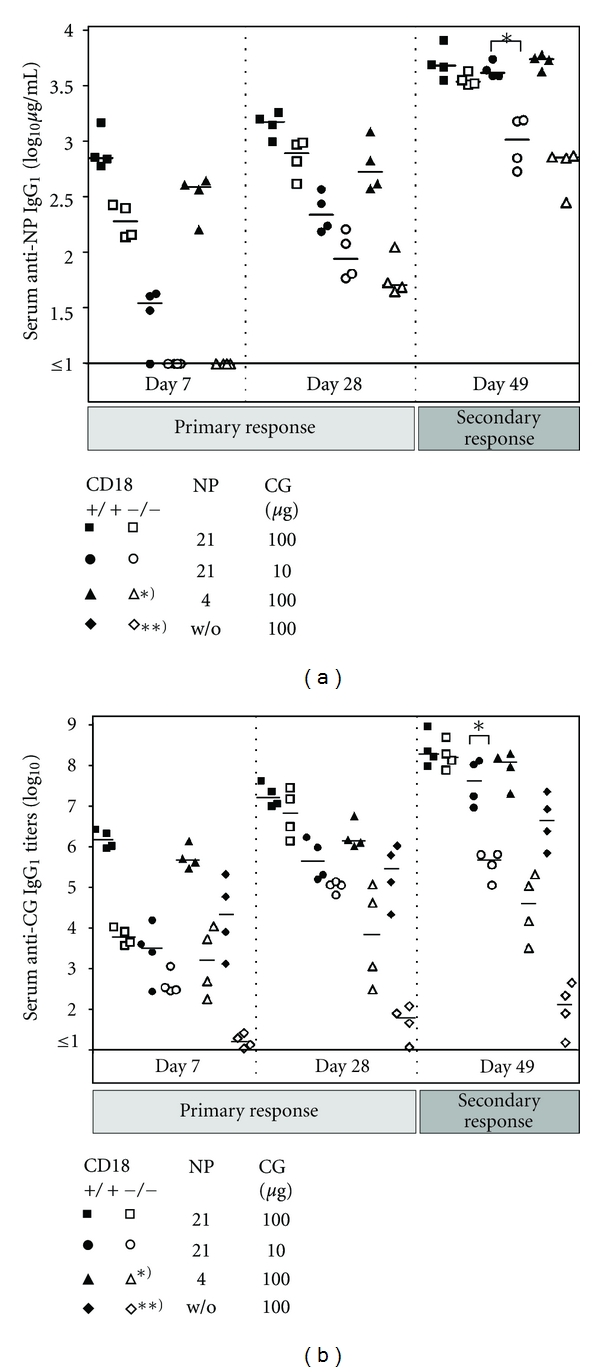
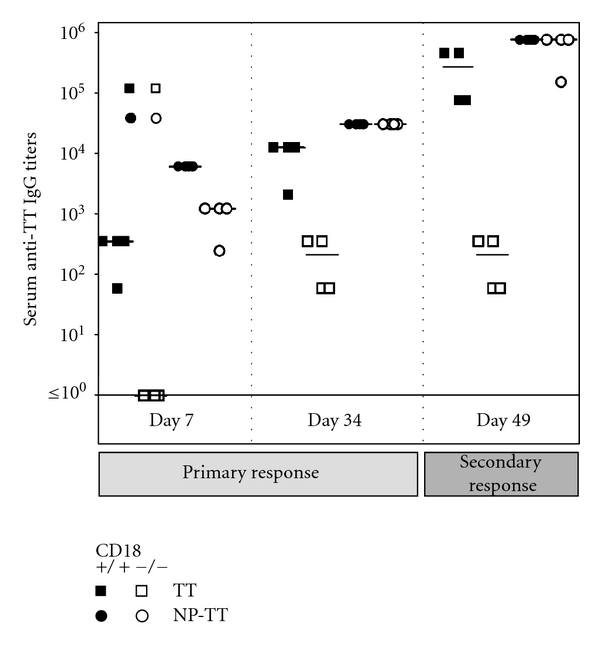
Absence of β₂ integrins (CD11/CD18) leads to leukocyte-adhesion deficiency-1 (LAD1), a rare primary immunodeficiency syndrome. Although extensive in vitro work has established an essential function of β₂ integrins in adhesive and signaling properties for cells of the innate and adaptive immune system, their respective participation in an altered adaptive immunity in LAD1 patients are complex and only partly understood in vivo. Therefore, we investigated adaptive immune responses towards different T-dependent antigens in a murine LAD1 model of β₂ integrin-deficiency (CD18⁻/⁻). CD18⁻/⁻ mice generated only weak IgG responses after immunization with tetanus toxoid (TT). In contrast, robust hapten- and protein-specific immune responses were observed after immunization with highly haptenated antigens such as (4-hydroxy-3-nitrophenyl)₂₁ acetyl chicken γ globulin (NP₂₁-CG), even though regularly structured germinal centers with specificity for the defined antigens/haptens in CD18⁻/⁻ mice remained absent. However, a decrease in the hapten/protein ratio lowered the efficacy of immune responses in CD18⁻/⁻ mice, whereas a mere reduction of the antigen dose was less crucial. Importantly, haptenation of TT with NP (NP-TT) efficiently restored a robust IgG response also to TT. Our findings may stimulate further studies on a modification of vaccination strategies using highly haptenated antigens in individuals suffering from LAD1.
Figures
References
-
- Balkow S, Heinz S, Schmidbauer P, et al. LFA-1 activity state on dendritic cells regulates contact duration with T cells and promotes T-cell priming. Blood. 2010;116(11):1885–1894. - PubMed
-
- Carrasco YR, Fleire SJ, Cameron T, Dustin ML, Batista FD. LFA-1/ICAM-1 interaction lowers the threshold of B cell activation by facilitating B cell adhesion and synapse formation. Immunity. 2004;20(5):589–599. - PubMed
-
- Grakoui A, Bromley SK, Sumen C, et al. The immunological synapse: a molecular machine controlling T cell activation. Science. 1999;285(5425):221–227. - PubMed
-
- Iezzi G, Karjalainen K, Lanzavecchia A. The duration of antigenic stimulation determines the fate of naive and effector T cells. Immunity. 1998;8(1):89–95. - PubMed
-
- Perez OD, Mitchell D, Jager GC, et al. Leukocyte functional antigen 1 lowers T cell activation thresholds and signaling through cytohesin-1 and Jun-activating binding protein 1. Nature Immunology. 2003;4(11):1083–1092. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
