

Vascular lumen formation
- PMID: 22474612
- PMCID: PMC3312398
- DOI: 10.1101/cshperspect.a006619
Vascular lumen formation
Abstract
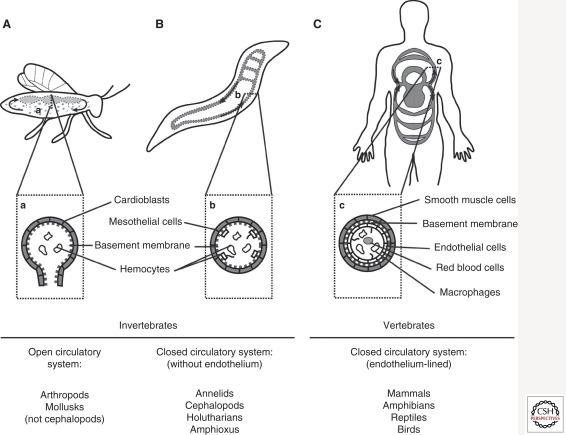
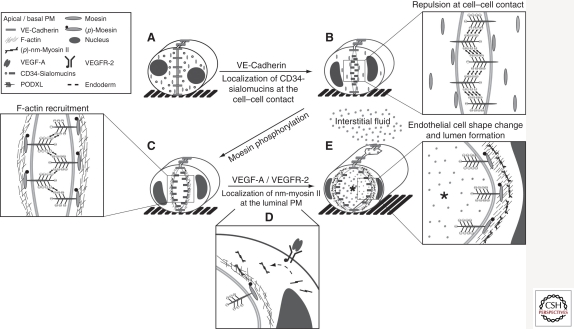
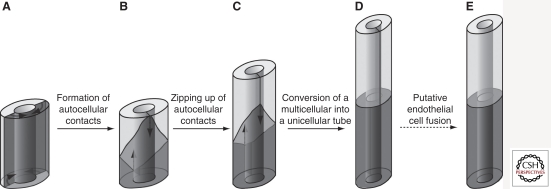
The vascular system developed early in evolution. It is required in large multicellular organisms for the transport of nutrients, oxygen, and waste products to and from tissues. The vascular system is composed of hollow tubes, which have a high level of complexity in vertebrates. Vasculogenesis describes the de novo formation of blood vessels, e.g., aorta formation in vertebrate embryogenesis. In contrast, angiogenesis is the formation of blood vessels from preexisting ones, e.g., sprouting of intersomitic blood vessels from the aorta. Importantly, the lumen of all blood vessels in vertebrates is lined and formed by endothelial cells. In both vasculogenesis and angiogenesis, lumen formation takes place in a cord of endothelial cells. It involves a complex molecular mechanism composed of endothelial cell repulsion at the cell-cell contacts within the endothelial cell cords, junctional rearrangement, and endothelial cell shape change. As the vascular system also participates in the course of many diseases, such as cancer, stroke, and myocardial infarction, it is important to understand and make use of the molecular mechanisms of blood vessel formation to better understand and manipulate the pathomechanisms involved.
Figures
References
-
- Adachi T, Tomita M, Yoshizato K 2005. Synthesis of prolyl 4-hydroxylase αa subunit and type IV collagen in hemocytic granular cells of silkworm, Bombyx mori: Involvement of type IV collagen in self-defense reaction and metamorphosis. Matrix Biol 24: 136–154 - PubMed
-
- Adams RH, Alitalo K 2007. Molecular regulation of angiogenesis and lymphangiogenesis. Nat Rev Mol Cell Biol 8: 464–478 - PubMed
-
- Barber VC, Graziadei P 1965. The fine structure of cephalopod blood vessels. I. Some smaller peripheral vessels. Z Zellforsch Mikrosk Anat 66: 765–781 - PubMed
-
- Bazzoni G, Dejana E 2004. Endothelial cell-to-cell junctions: Molecular organization and role in vascular homeostasis. Physiol Rev 84: 869–901 - PubMed
-
- Betsholtz C, Lindblom P, Gerhardt H 2005. Role of pericytes in vascular morphogenesis. EXS 115–125 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials