

Evolutionary development of the plant and spore wall
- PMID: 22476497
- PMCID: PMC3220415
- DOI: 10.1093/aobpla/plr027
Evolutionary development of the plant and spore wall
Abstract
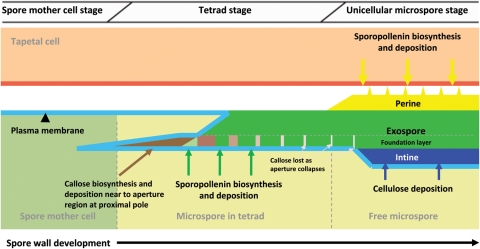
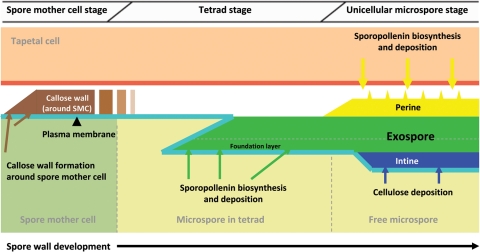
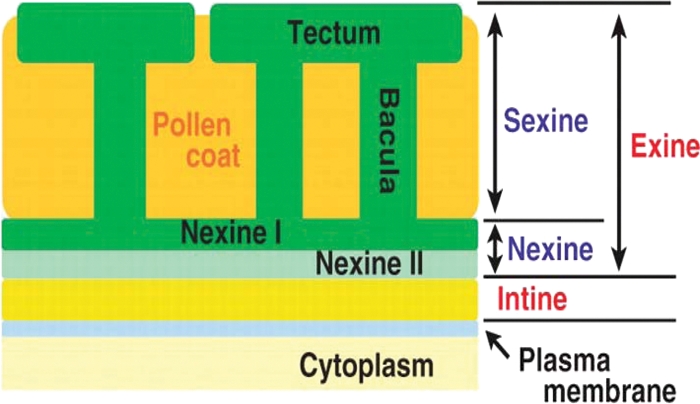
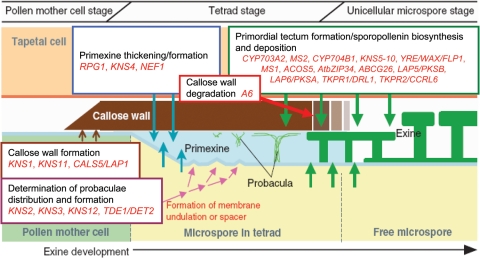
Background and aims: Many key innovations were required to enable plants to colonize terrestrial habitats successfully. One of these was the acquisition of a durable spore/pollen wall capable of withstanding the harsh desiccating and UV-B-rich environment encountered on land. The spores of 'lower' spore-bearing plants and the pollen of 'higher' seed plants are homologous. In recent years, researchers have begun to investigate the molecular genetics of pollen wall development in angiosperms (including the model organism Arabidopsis thaliana). However, research into the molecular genetics of spore wall development in more basal plants has thus far been extremely limited. This review summarizes the literature on spore/pollen wall development, including the molecular genetics associated with pollen wall development in angiosperms, in a preliminary attempt to identify possible candidate genes involved in spore wall development in more basal plants.
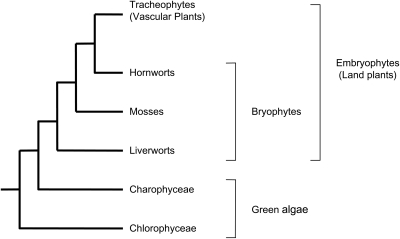
Presence in moss of genes involved in pollen wall development: Bioinformatic studies have suggested that genes implicated in pollen wall development in angiosperms are also present in moss and lycopsids, and may therefore be involved in spore wall development in basal plants. This suggests that the molecular genetics of spore/pollen development are highly conserved, despite the large morphological and functional differences between spores and pollen.
Future work: The use of high-throughput sequencing strategies and/or microarray experiments at an appropriate stage of 'lower' land plant sporogenesis will allow the identification of candidate genes likely to be involved in the development of the spore wall by way of comparison with those genes known to be involved in pollen wall development. Additionally, by conducting gene knock-out and gene swap experiments between 'lower' land plant species, such as the moss model species Physcomitrella patens, and the angiosperm model species arabidopsis it will be possible to test the role of these candidate genes.
Figures
References
-
- Aarts MGM, Hodge R, Halantidis K, Florack D, Wilson AZ, Mulligan BJ, Stiekema WJ, Scott R, Pereira A. The Arabidopsis MALE STERILITY 2 protein shares similarity with reductases in elongation/condensation complexes. The Plant Journal. 1997;12:615–623. - PubMed
-
- Ahlers F, Thom I, Lambert J, Kuckuk R, Wiermann R. 1H NMR analysis of sporopollenin from Typha angustifolia. Phytochemistry. 1999;5:1095–1098.
-
- Ariizumi T, Hatakeyama K, Hinata K, Sato S, Kato T, Tabata S, Toriyama K. A novel male-sterile mutant of Arabidopsis thaliana, faceless pollen-1, produces pollen with a smooth surface and an acetolysis-sensitive exine. Plant Molecular Biology. 2003;53:107–116. - PubMed
-
- Ariizumi T, Hatakeyama K, Hinata K, Inatsugi R, Nishida I, Sato S, Kato T, Tabata S, Toriyama K. Disruption of the novel plant protein NEF1 affects lipid accumulation in the plastids of the tapetum and exine formation of pollen, resulting in male sterility in Arabidopsis thaliana. The Plant Journal. 2004;39:170–181. - PubMed
-
- Ariizumi T, Hatakeyama K, Hinata K, Sato S, Kato T, Tabata S, Toriyama K. The HKM gene, which is identical to the MS1 gene of Arabidopsis thaliana, is essential for primexine formation and exine pattern formation. Sexual Plant Reproduction. 2005;18:1–7.
LinkOut - more resources
Full Text Sources
Research Materials
