

Mammalian neurogenesis requires Treacle-Plk1 for precise control of spindle orientation, mitotic progression, and maintenance of neural progenitor cells
- PMID: 22479190
- PMCID: PMC3315461
- DOI: 10.1371/journal.pgen.1002566
Mammalian neurogenesis requires Treacle-Plk1 for precise control of spindle orientation, mitotic progression, and maintenance of neural progenitor cells
Abstract
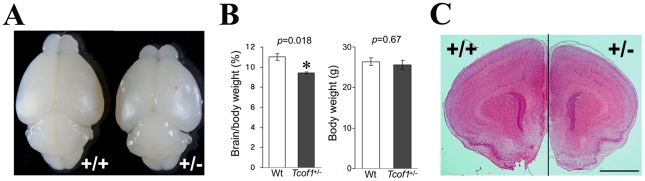
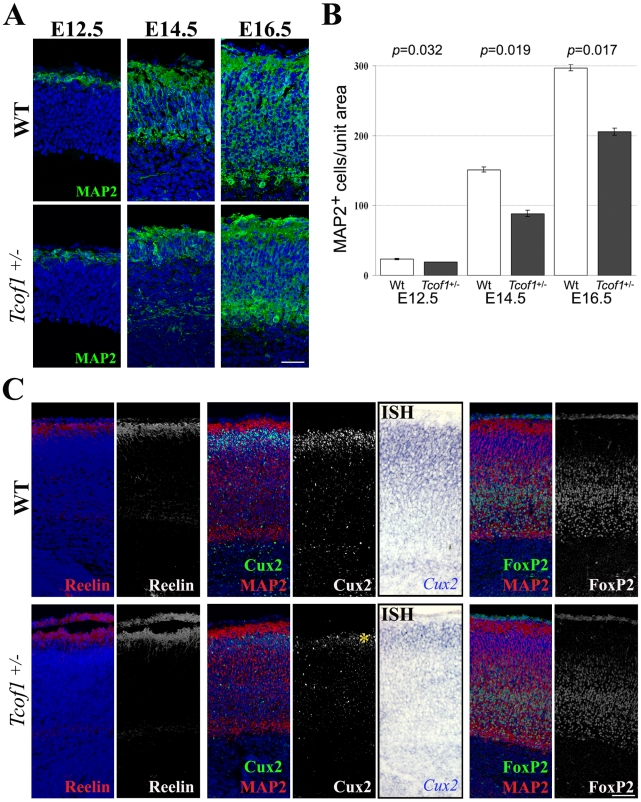
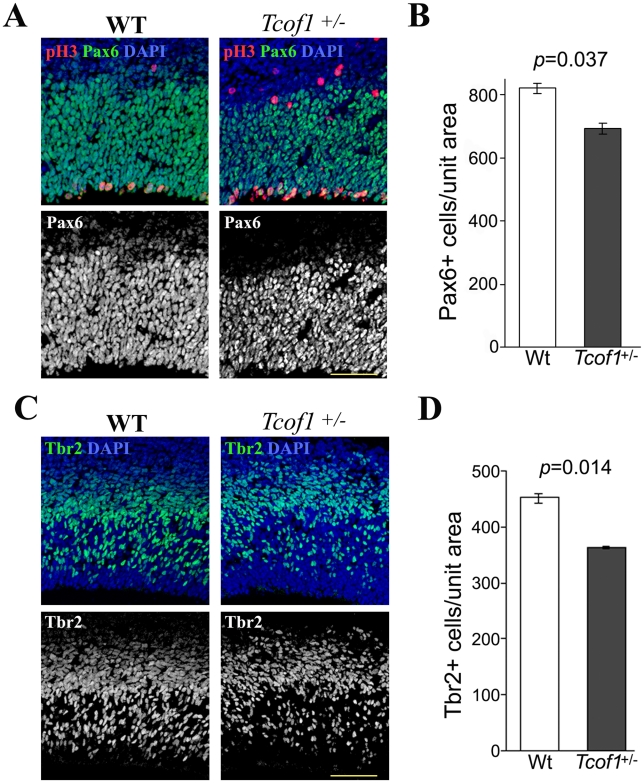
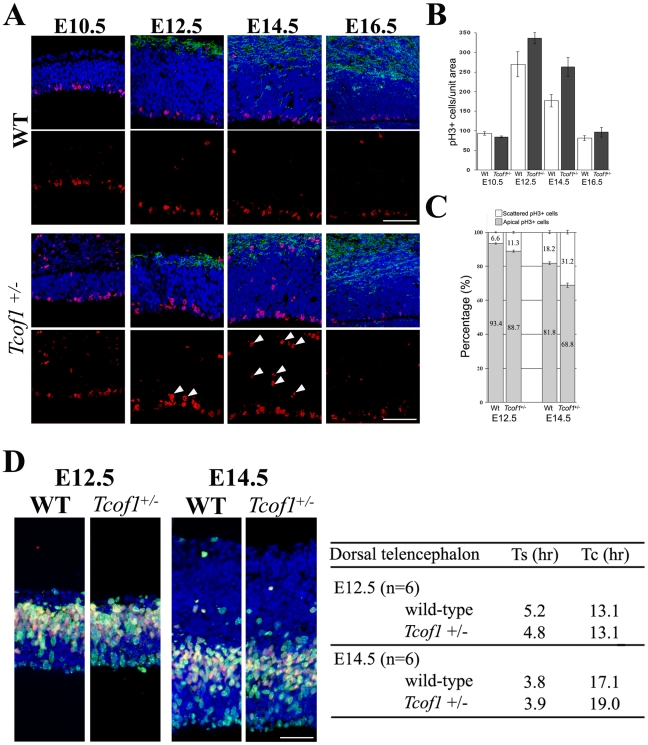
The cerebral cortex is a specialized region of the brain that processes cognitive, motor, somatosensory, auditory, and visual functions. Its characteristic architecture and size is dependent upon the number of neurons generated during embryogenesis and has been postulated to be governed by symmetric versus asymmetric cell divisions, which mediate the balance between progenitor cell maintenance and neuron differentiation, respectively. The mechanistic importance of spindle orientation remains controversial, hence there is considerable interest in understanding how neural progenitor cell mitosis is controlled during neurogenesis. We discovered that Treacle, which is encoded by the Tcof1 gene, is a novel centrosome- and kinetochore-associated protein that is critical for spindle fidelity and mitotic progression. Tcof1/Treacle loss-of-function disrupts spindle orientation and cell cycle progression, which perturbs the maintenance, proliferation, and localization of neural progenitors during cortical neurogenesis. Consistent with this, Tcof1(+/-) mice exhibit reduced brain size as a consequence of defects in neural progenitor maintenance. We determined that Treacle elicits its effect via a direct interaction with Polo-like kinase1 (Plk1), and furthermore we discovered novel in vivo roles for Plk1 in governing mitotic progression and spindle orientation in the developing mammalian cortex. Increased asymmetric cell division, however, did not promote increased neuronal differentiation. Collectively our research has therefore identified Treacle and Plk1 as novel in vivo regulators of spindle fidelity, mitotic progression, and proliferation in the maintenance and localization of neural progenitor cells. Together, Treacle and Plk1 are critically required for proper cortical neurogenesis, which has important implications in the regulation of mammalian brain size and the pathogenesis of congenital neurodevelopmental disorders such as microcephaly.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
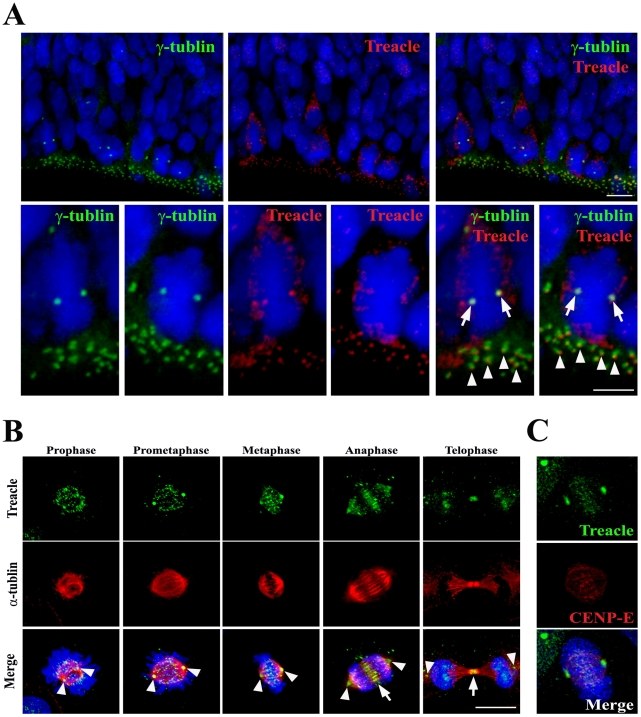
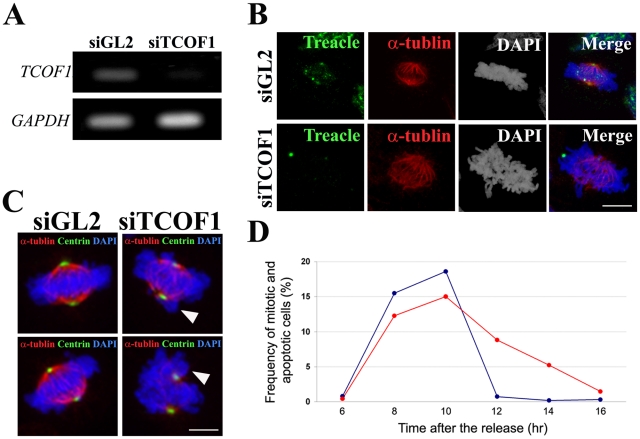
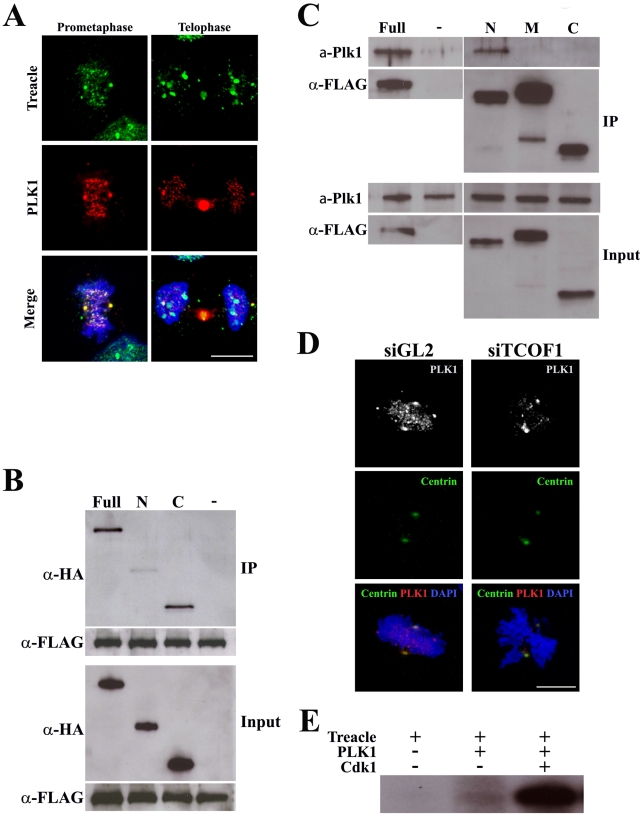
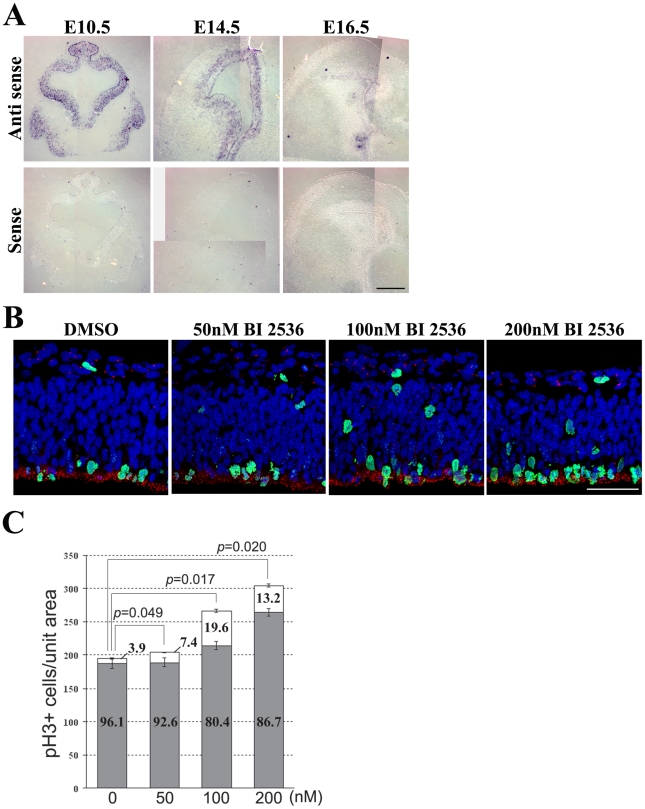
References
-
- Noctor SC, Flint AC, Weissman TA, Dammerman RS, Kriegstein AR. Neurons derived from radial glial cells establish radial units in neocortex. Nature. 2001;409:714–720. - PubMed
-
- Miyata T, Kawaguchi A, Saito K, Kawano M, Muto T, et al. Asymmetric production of surface-dividing and non-surface-dividing cortical progenitor cells. Development. 2004;131:3133–3145. - PubMed
-
- Noctor SC, Martinez-Cerdeno V, Ivic L, Kriegstein AR. Cortical neurons arise in symmetric and asymmetric division zones and migrate through specific phases. Nat Neurosci. 2004;7:136–144. - PubMed
-
- Chenn A, McConnell SK. Cleavage orientation and the asymmetric inheritance of Notch1 immunoreactivity in mammalian neurogenesis. Cell. 1995;82:631–641. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
