

Carbohydrate Derived Fulvic Acid: An in vitro Investigation of a Novel Membrane Active Antiseptic Agent Against Candida albicans Biofilms
- PMID: 22479260
- PMCID: PMC3314872
- DOI: 10.3389/fmicb.2012.00116
Carbohydrate Derived Fulvic Acid: An in vitro Investigation of a Novel Membrane Active Antiseptic Agent Against Candida albicans Biofilms
Abstract
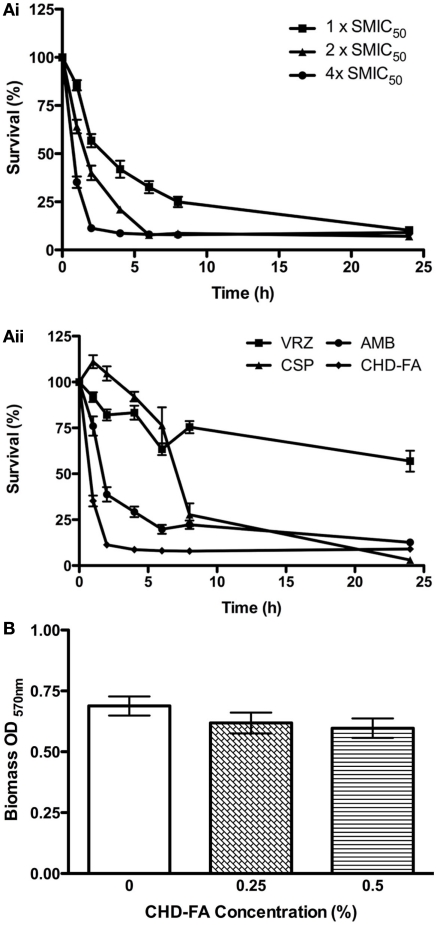
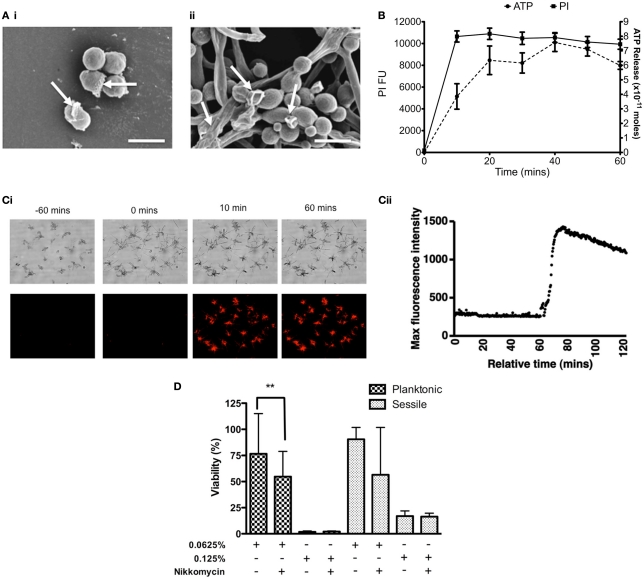
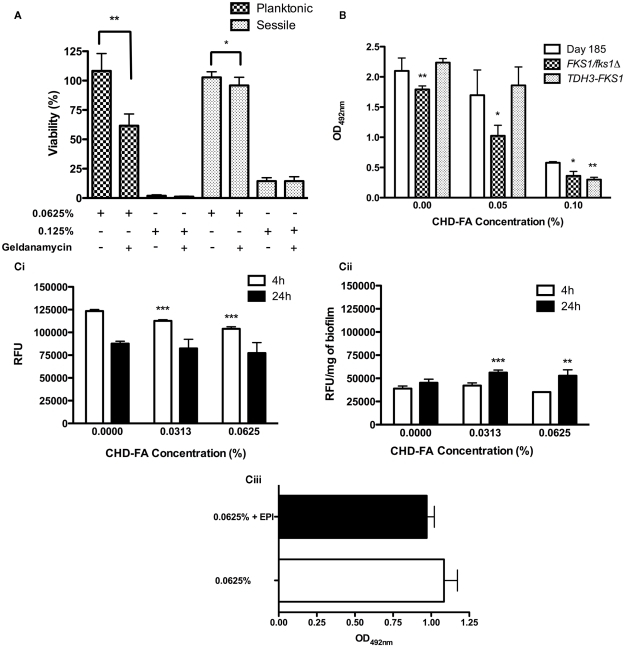
Carbohydrate derived fulvic acid (CHD-FA) is a heat stable low molecular weight, water soluble, cationic, colloidal material with proposed therapeutic properties. The aim of this study was to evaluate the antifungal activity of CHD-FA against Candida albicans, and to characterize its mode of action. A panel of C. albicans isolates (n = 50) derived from a range of clinical specimens were grown planktonically and as biofilms, and the minimum inhibitory concentrations determined. Scanning electron microscopy was performed to examine ultrastructural changes and different cell membrane assays were used to determine its mode of action. In addition, the role of C. albicans biofilm resistance mechanisms were investigated to determine their effects on CHD-FA activity. CHD-FA was active against planktonic and sessile C. albicans at concentrations 0.125 and 0.25% respectively, and was shown to be fungicidal, acting through disruption of the cell membrane activity. Resistance mechanisms, including matrix, efflux, and stress, had a limited role upon CHD-FA activity. Overall, based on the promising in vitro spectrum of activity and minimal biofilm resistance of the natural and cheap antiseptic CHD-FA, further studies are required to determine its applicability for clinical use.
Keywords: Candida albicans; antiseptic; biofilm; fulvic acid.
Figures
References
-
- Clinical Laboratory Standards Institute (CLSI) (2008). Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeast, Approved Standard – 3rd Edn, CLSI document M27-A3. Wayne, PA: CLSI
LinkOut - more resources
Full Text Sources