

The conserved Cockayne syndrome B-piggyBac fusion protein (CSB-PGBD3) affects DNA repair and induces both interferon-like and innate antiviral responses in CSB-null cells
- PMID: 22483866
- PMCID: PMC3340519
- DOI: 10.1016/j.dnarep.2012.02.004
The conserved Cockayne syndrome B-piggyBac fusion protein (CSB-PGBD3) affects DNA repair and induces both interferon-like and innate antiviral responses in CSB-null cells
Abstract
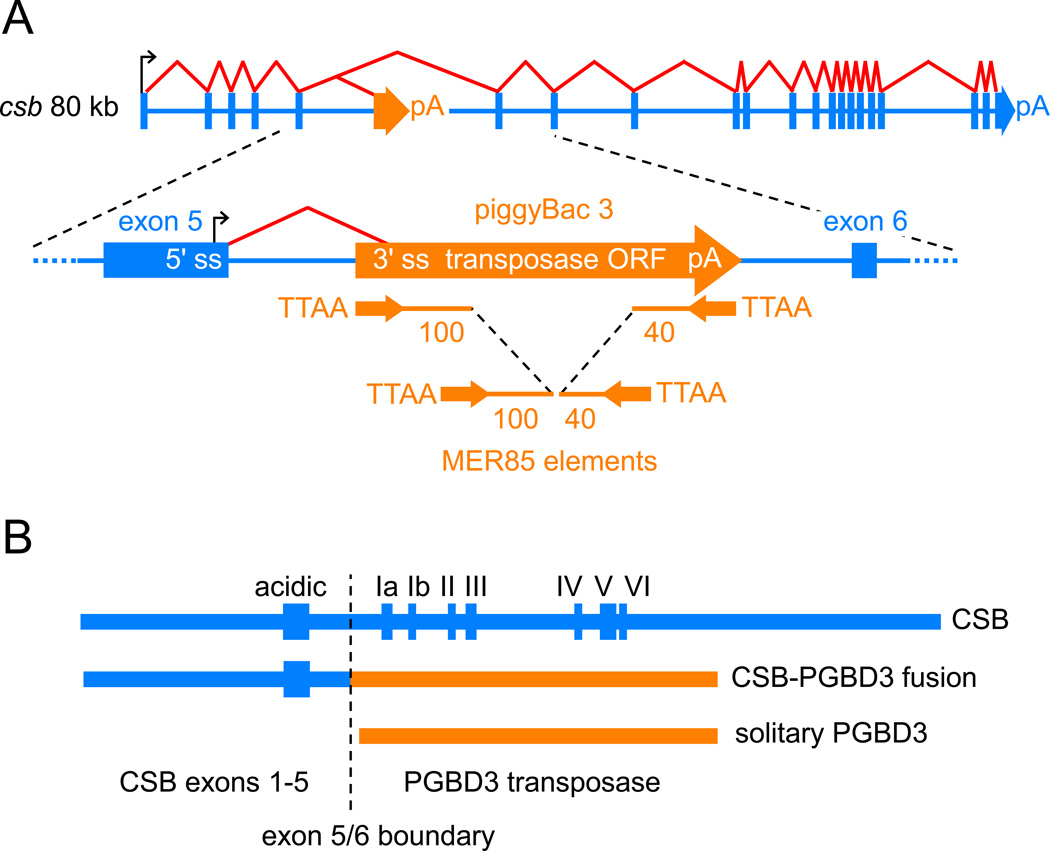
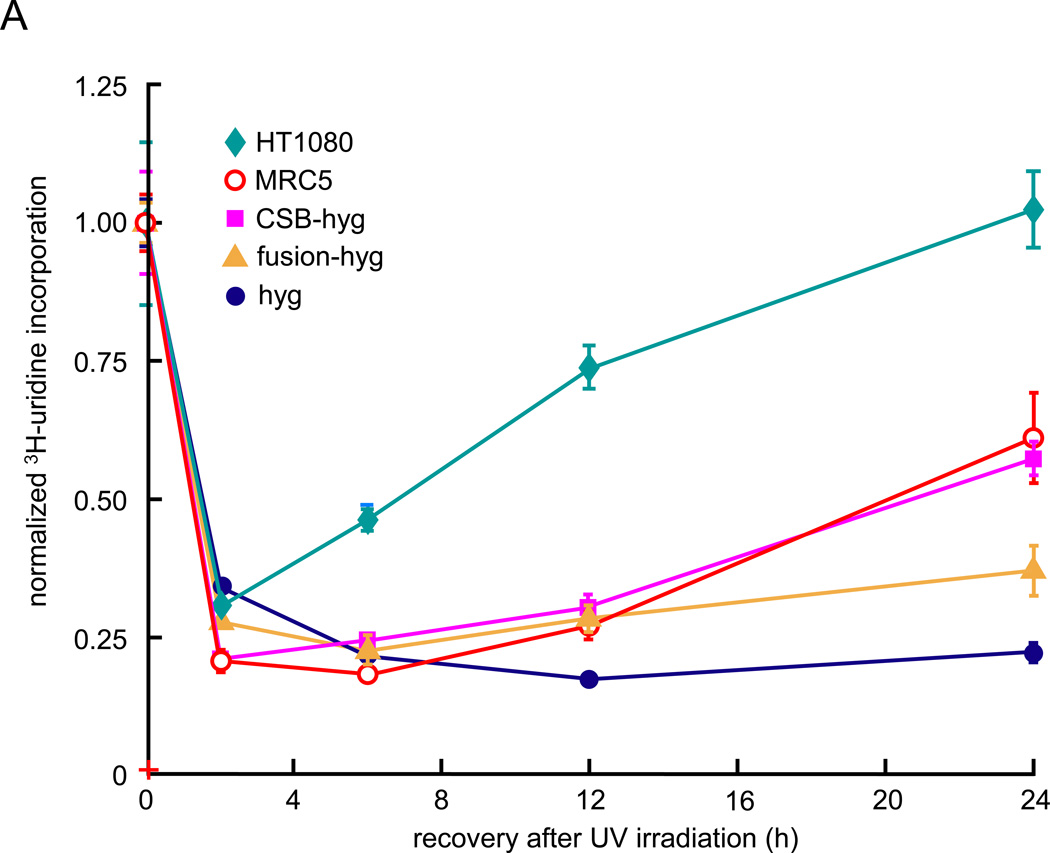
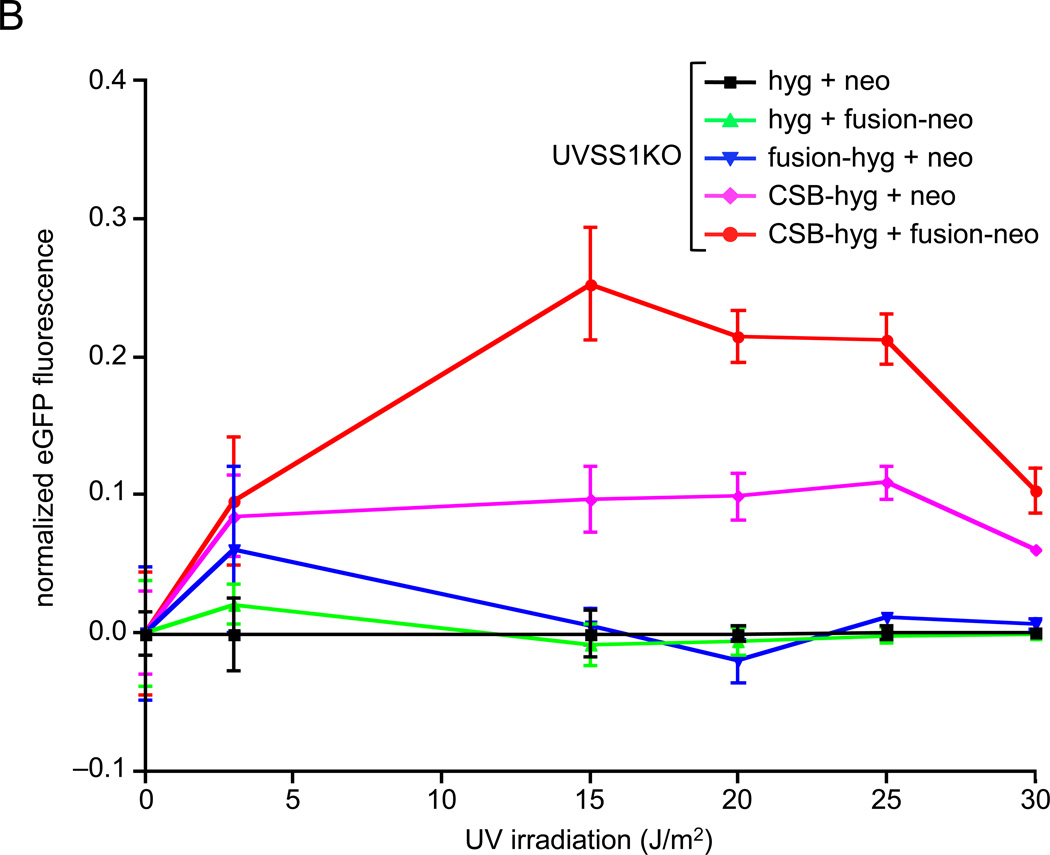
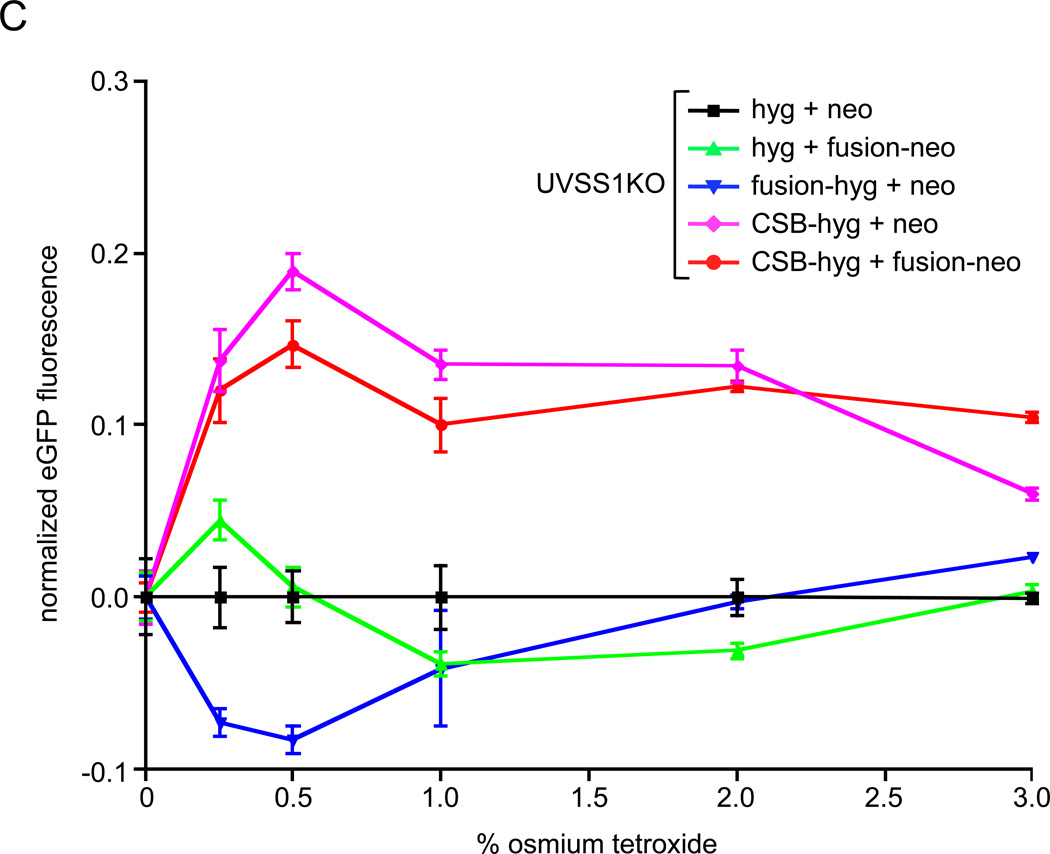
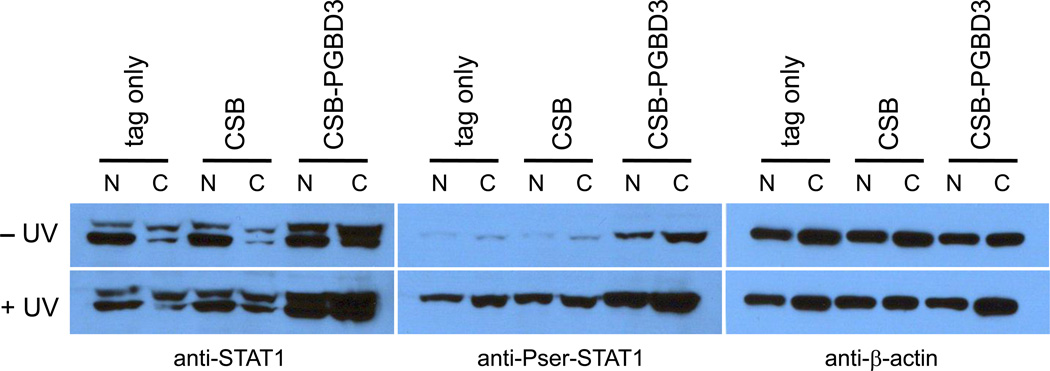
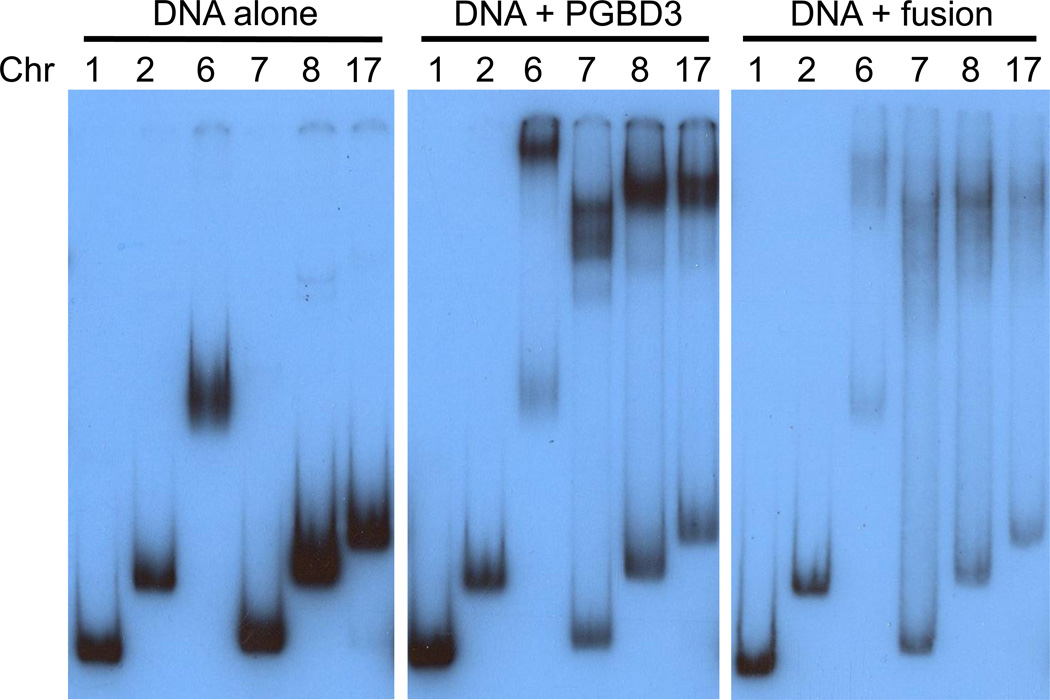
Cockayne syndrome is a segmental progeria most often caused by mutations in the CSB gene encoding a SWI/SNF-like ATPase required for transcription-coupled DNA repair (TCR). Over 43Mya before marmosets diverged from humans, a piggyBac3 (PGBD3) transposable element integrated into intron 5 of the CSB gene. As a result, primate CSB genes now generate both CSB protein and a conserved CSB-PGBD3 fusion protein in which the first 5 exons of CSB are alternatively spliced to the PGBD3 transposase. Using a host cell reactivation assay, we show that the fusion protein inhibits TCR of oxidative damage but facilitates TCR of UV damage. We also show by microarray analysis that expression of the fusion protein alone in CSB-null UV-sensitive syndrome (UVSS) cells induces an interferon-like response that resembles both the innate antiviral response and the prolonged interferon response normally maintained by unphosphorylated STAT1 (U-STAT1); moreover, as might be expected based on conservation of the fusion protein, this potentially cytotoxic interferon-like response is largely reversed by coexpression of functional CSB protein. Interestingly, expression of CSB and the CSB-PGBD3 fusion protein together, but neither alone, upregulates the insulin growth factor binding protein IGFBP5 and downregulates IGFBP7, suggesting that the fusion protein may also confer a metabolic advantage, perhaps in the presence of DNA damage. Finally, we show that the fusion protein binds in vitro to members of a dispersed family of 900 internally deleted piggyBac elements known as MER85s, providing a potential mechanism by which the fusion protein could exert widespread effects on gene expression. Our data suggest that the CSB-PGBD3 fusion protein is important in both health and disease, and could play a role in Cockayne syndrome.
Copyright © 2012 Elsevier B.V. All rights reserved.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Figures
References
-
- Beerens N, Hoeijmakers JH, Kanaar R, Vermeulen W, Wyman C. The CSB protein actively wraps DNA. J Biol Chem. 2005;280:4722–4729. - PubMed
-
- Nardo T, Oneda R, Spivak G, Vaz B, Mortier L, Thomas P, Orioli D, Laugel V, Stary A, Hanawalt PC, Sarasin A, Stefanini M. A UV-sensitive syndrome patient with a specific CSA mutation reveals separable roles for CSA in response to UV and oxidative DNA damage. Proc Natl Acad Sci U S A. 2009;106:6209–6214. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
