

EB1-recruited microtubule +TIP complexes coordinate protrusion dynamics during 3D epithelial remodeling
- PMID: 22483942
- PMCID: PMC3350573
- DOI: 10.1016/j.cub.2012.02.069
EB1-recruited microtubule +TIP complexes coordinate protrusion dynamics during 3D epithelial remodeling
Abstract
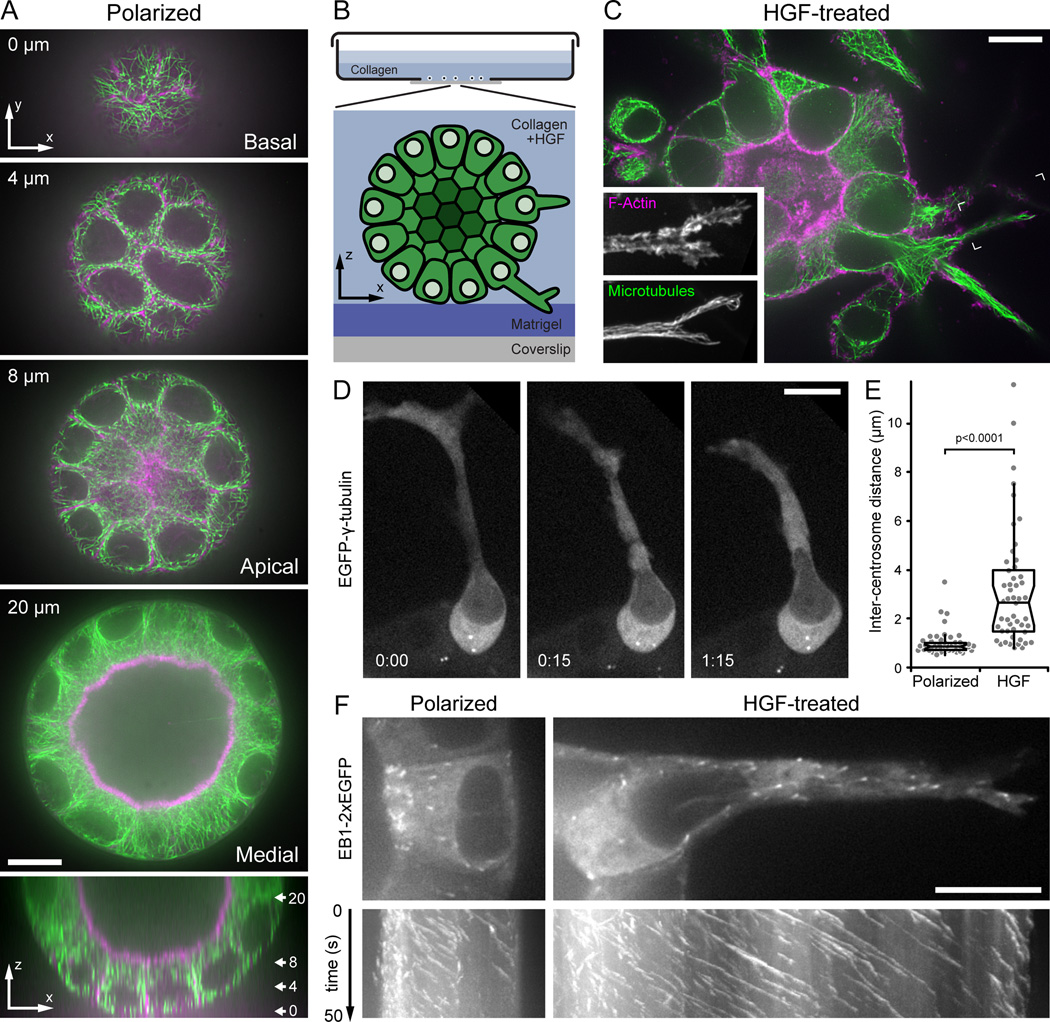
Background: Epithelial remodeling, in which apical-basal polarized cells switch to a migratory phenotype, plays a central role in development and disease of multicellular organisms. Although dynamic microtubules (MTs) are required for directed migration on flat surfaces, how MT dynamics are controlled or contribute to epithelial remodeling in a more physiological three-dimensional (3D) environment is not understood. We use confocal live-cell imaging to analyze MT function and dynamics during 3D epithelial morphogenesis and remodeling of polarized Madin-Darby canine kidney epithelial cells that undergo partial epithelial-to-mesenchymal transition in response to hepatocyte growth factor (HGF).
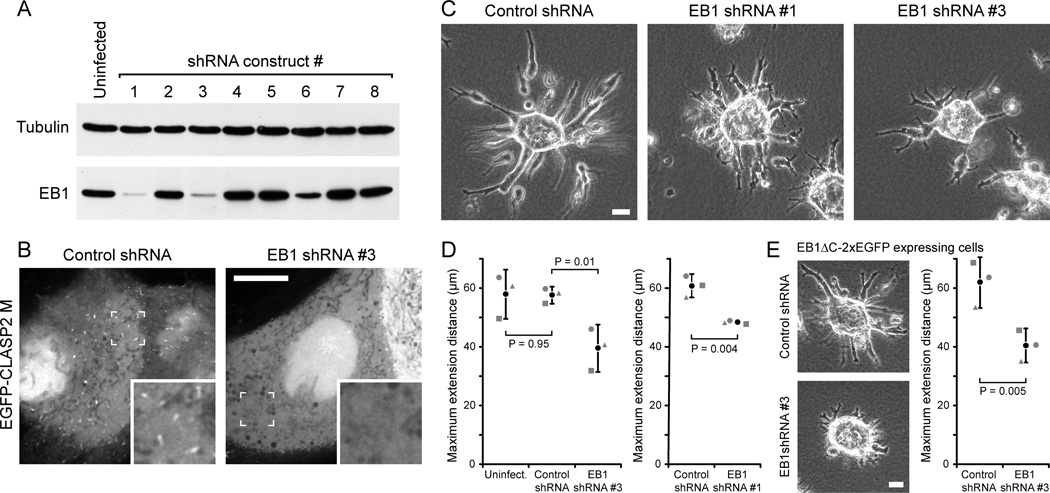
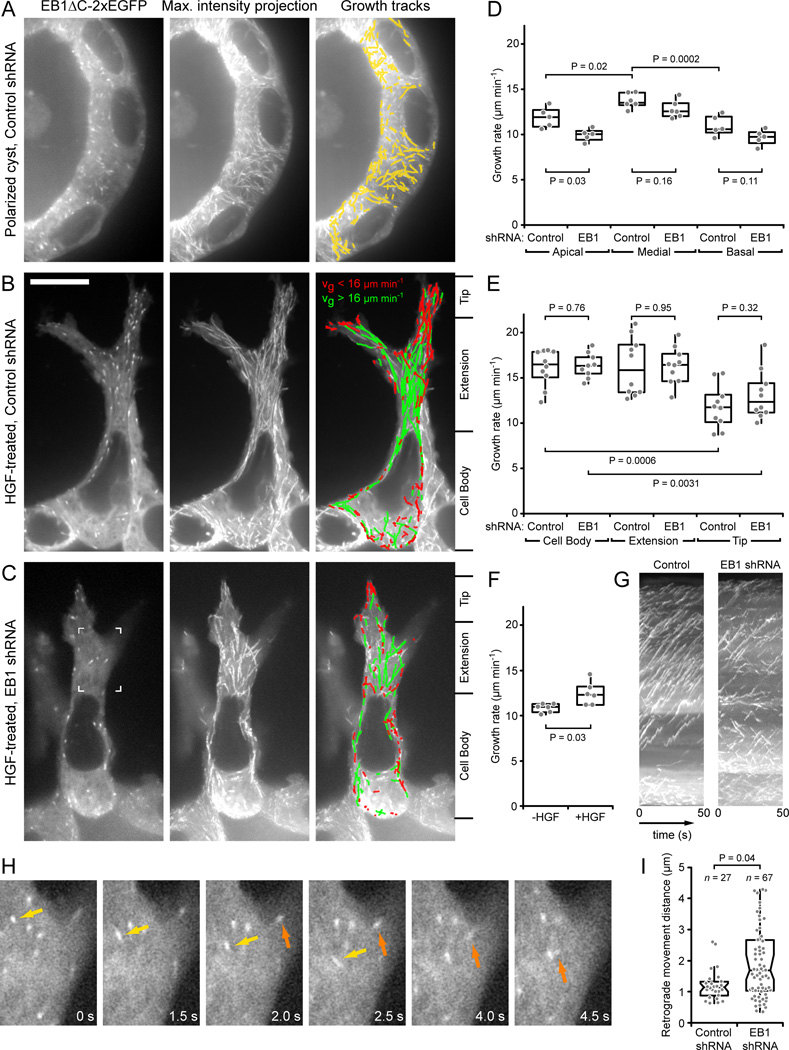
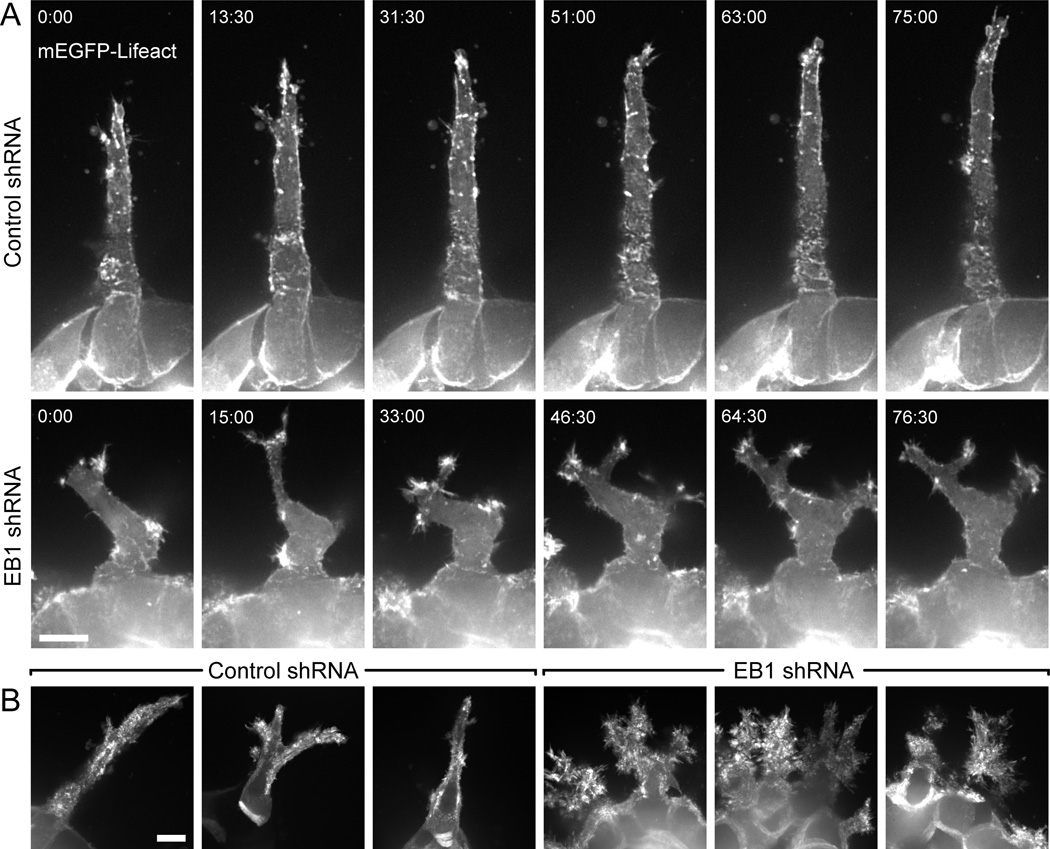
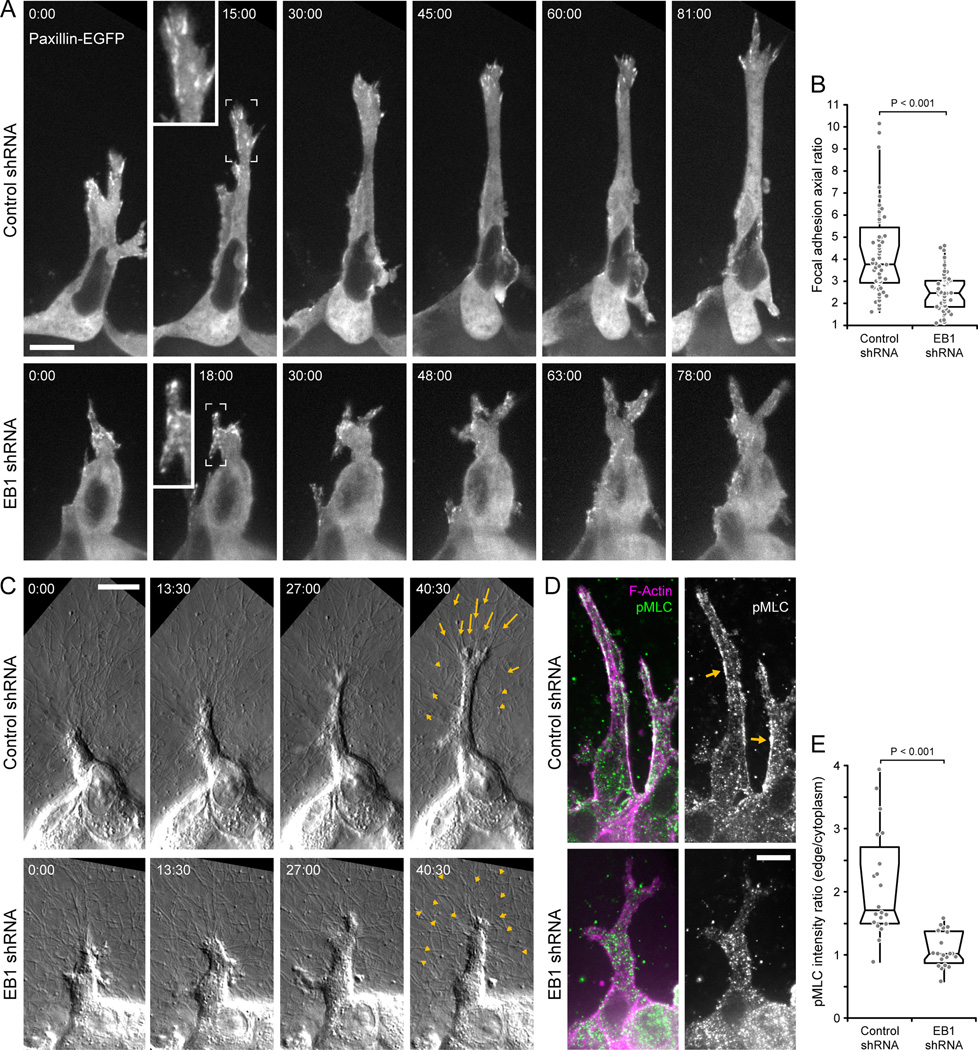
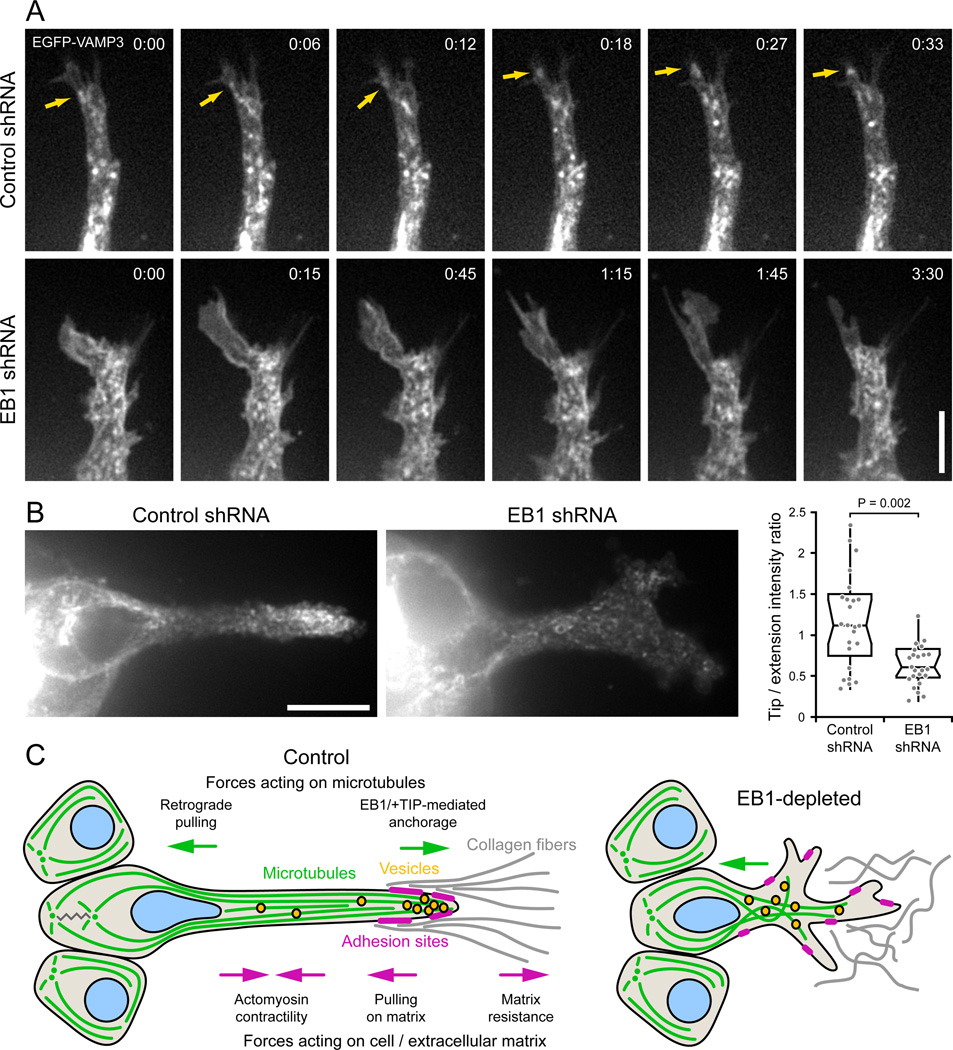
Results: We find that HGF treatment increases MT growth rate before morphological changes are evident and that large numbers of MTs grow into HGF-induced cell extensions independent of centrosome reorientation. Using lentivirus-mediated small hairpin RNA, we demonstrate that EB1, an adaptor protein that mediates recruitment of numerous other +TIP proteins to growing MT plus ends, is required for this HGF-induced MT reorganization. We further show that protrusion and adhesion dynamics are disorganized and that vesicular trafficking to the tip of HGF-induced cell extensions is disrupted in EB1-depleted cells.
Conclusions: We conclude that EB1-mediated interactions with growing MTs are important to coordinate cell shape changes and directed migration into the surrounding extracellular matrix during epithelial remodeling in a physiological 3D environment. In contrast, EB1 is not required for the establishment or maintenance of apical-basal cell polarity, suggesting different functions of +TIPs and MTs in different types of cell polarity.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
