

BDMC33, A curcumin derivative suppresses inflammatory responses in macrophage-like cellular system: role of inhibition in NF-κB and MAPK signaling pathways
- PMID: 22489138
- PMCID: PMC3317699
- DOI: 10.3390/ijms13032985
BDMC33, A curcumin derivative suppresses inflammatory responses in macrophage-like cellular system: role of inhibition in NF-κB and MAPK signaling pathways
Abstract
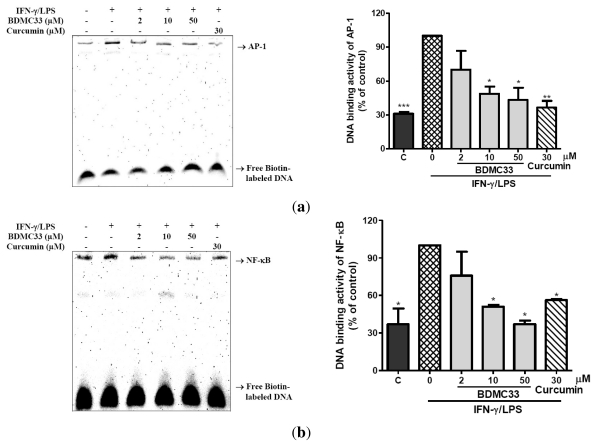
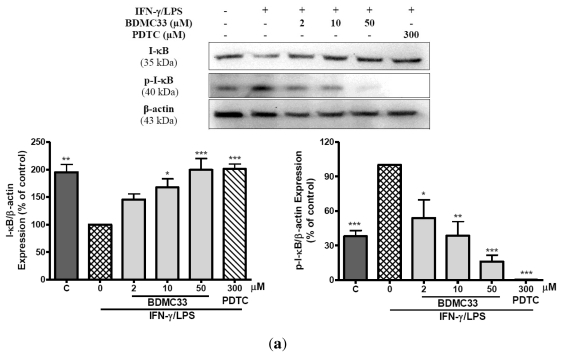
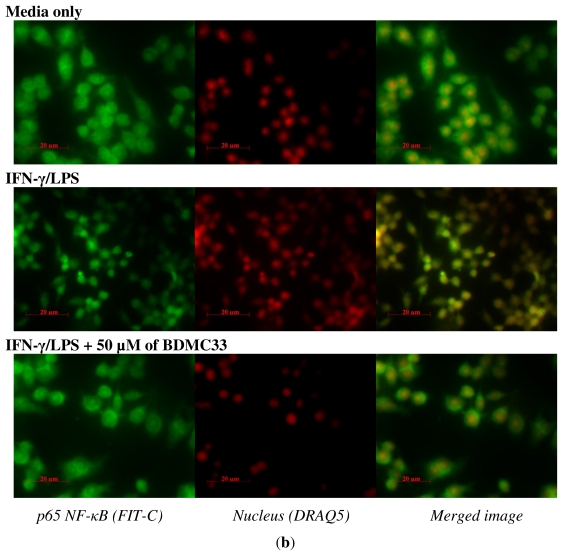
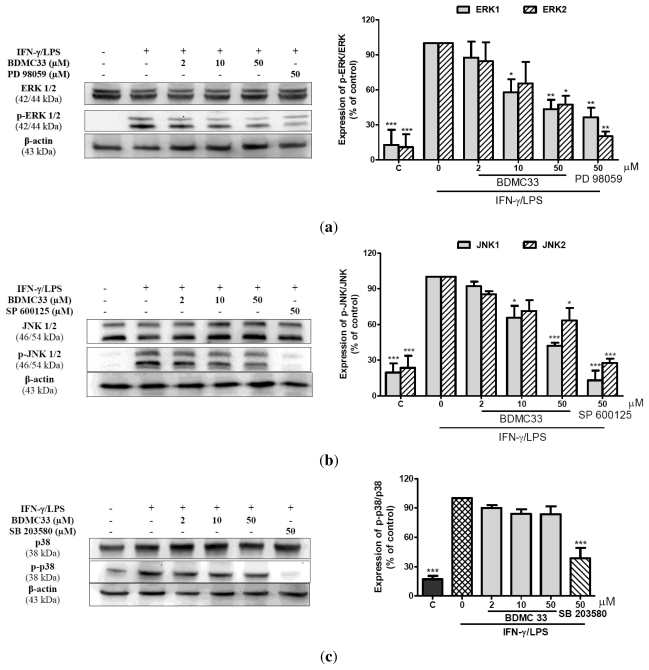
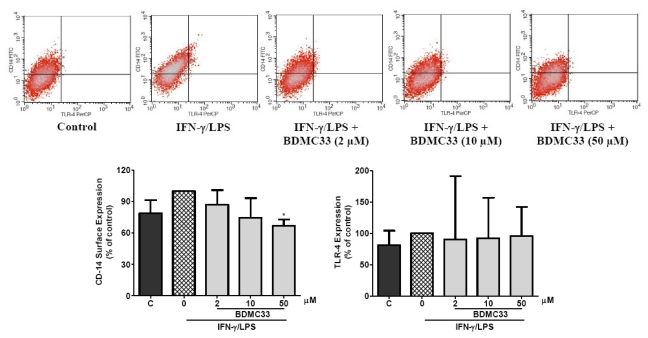
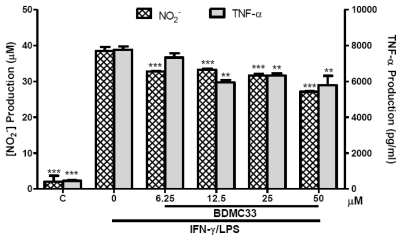
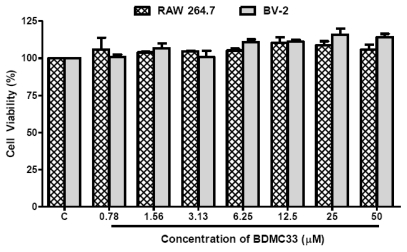
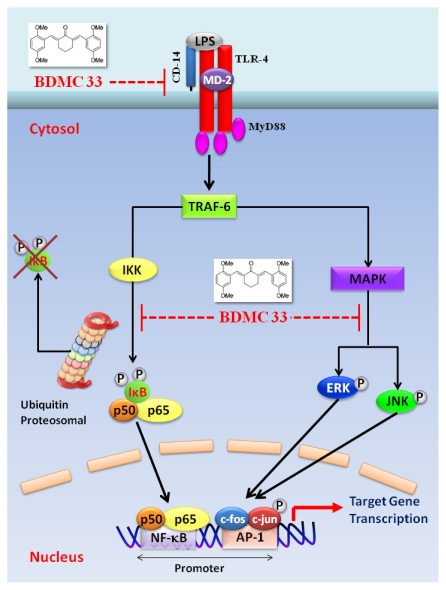
Our preliminary screening has shown that curcumin derivative BDMC33 [2,6-bis(2,5-dimethoxybenzylidene)cyclohexanone] exerted promising nitric oxide inhibitory activity in activated macrophages. However, the molecular basis and mechanism for its pharmacological action is yet to be elucidated. The aim of this study was to investigate the anti-inflammatory properties of BDMC33 and elucidate its underlying mechanism action in macrophage cells. Our current study demonstrated that BDMC33 inhibits the secretion of major pro-inflammatory mediators in stimulated macrophages, and includes NO, TNF-α and IL-1β through interference in both nuclear factor kappaB (NF-κB) and mitogen activator protein kinase (MAPK) signaling cascade in IFN-γ/LPS-stimulated macrophages. Moreover, BDMC33 also interrupted LPS signaling through inhibiting the surface expression of CD-14 accessory molecules. In addition, the inhibitory action of BDMC33 not only restricted the macrophages cell (RAW264.7), but also inhibited the secretion of NO and TNF-α in IFN-γ/LPS-challenged microglial cells (BV-2). The experimental data suggests the inflammatory action of BDMC33 on activated macrophage-like cellular systems, which could be used as a future therapeutic agent in the management of chronic inflammatory diseases.
Keywords: BV-2; MAPK; NF-κB; RAW 264.7; anti-inflammatory; curcumin.
Figures
References
-
- Rodriguez-Vita J., Lawrence T. The resolution of inflammation and cancer. Cytokine Growth Factor Rev. 2010;2:61–65. - PubMed
-
- Schwacha M.G. Macrophages and post-burn immune dysfunction. Burns. 2003;29:1–14. - PubMed
-
- Bowdish D.M.E., Loffredo M.S., Mukhopadhyay S., Mantovani A., Gordon S. Macrophage receptors implicated in the “adaptive” form of innate immunity. Microbes Infect. 2007;9:1680–1687. - PubMed
-
- Chan E.D., Riches D.W.H. IFN-γ + LPS induction of iNOS is modulated by ERK, JNK/SAPK, and p38mapk in a mouse macrophage cell line. Am. J. Physiol. Cell Physiol. 2001;280:441–450. - PubMed
-
- Held T.K., WeiHua X., Yuan L., Kalvakolanu D.V., Cross A.S. Gamma interferon uugments macrophage activation by lipopolysaccharide by two distinct mechanisms, at the signal transduction level and via an autocrine mechanism involving tumor necrosis factor alpha and interleukin-1. Infect. Immun. 1999;67:206–212. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
