

The role of muscle spindles in the development of the monosynaptic stretch reflex
- PMID: 22490553
- PMCID: PMC3434619
- DOI: 10.1152/jn.00074.2012
The role of muscle spindles in the development of the monosynaptic stretch reflex
Abstract
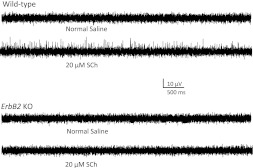
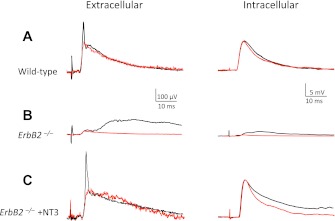
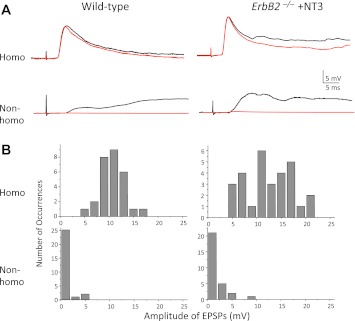
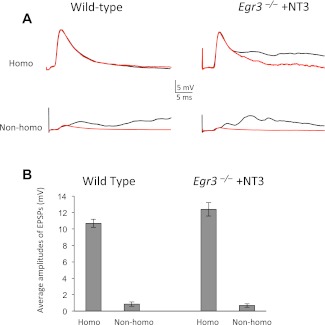
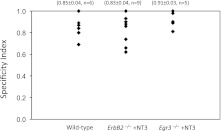
Muscle sensory axons induce the development of specialized intrafusal muscle fibers in muscle spindles during development, but the role that the intrafusal fibers may play in the development of the central projections of these Ia sensory axons is unclear. In the present study, we assessed the influence of intrafusal fibers in muscle spindles on the formation of monosynaptic connections between Ia (muscle spindle) sensory axons and motoneurons (MNs) using two transgenic strains of mice. Deletion of the ErbB2 receptor from developing myotubes disrupts the formation of intrafusal muscle fibers and causes a nearly complete absence of functional synaptic connections between Ia axons and MNs. Monosynaptic connectivity can be fully restored by postnatal administration of neurotrophin-3 (NT-3), and the synaptic connections in NT-3-treated mice are as specific as in wild-type mice. Deletion of the Egr3 transcription factor also impairs the development of intrafusal muscle fibers and disrupts synaptic connectivity between Ia axons and MNs. Postnatal injections of NT-3 restore the normal strengths and specificity of Ia-motoneuronal connections in these mice as well. Severe deficits in intrafusal fiber development, therefore, do not disrupt the establishment of normal, selective patterns of connections between Ia axons and MNs, although these connections require the presence of NT-3, normally supplied by intrafusal fibers, to be functional.
Figures
References
-
- Arber S, Ladle DR, Lin JH, Frank E, Jessell TM. ETS gene Er81 controls the formation of functional connections between group Ia sensory afferents and motor neurons. Cell 101: 485–498, 2000 - PubMed
-
- Boyd IA. Intrafusal muscle fibres in the cat and their motor control. In: Feedback and Motor Control in Invertebrates and Vertebrates, edited by Barnes WJP, Gladden MH. London: Croom Helm, 1985, p. 123–144
-
- Carr VM, Simpson SB., Jr Proliferative and degenerative events in the early development of chick dorsal root ganglia. I. Normal development. J Comp Neurol 182: 727–739, 1978 - PubMed
-
- Chen HH, Hippenmeyer S, Arber S, Frank E. Development of the monosynaptic stretch reflex circuit. Curr Opin Neurobiol 13: 96–102, 2003 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous