

Endogenous erythropoietin signaling facilitates skeletal muscle repair and recovery following pharmacologically induced damage
- PMID: 22490927
- PMCID: PMC3382092
- DOI: 10.1096/fj.11-196618
Endogenous erythropoietin signaling facilitates skeletal muscle repair and recovery following pharmacologically induced damage
Abstract
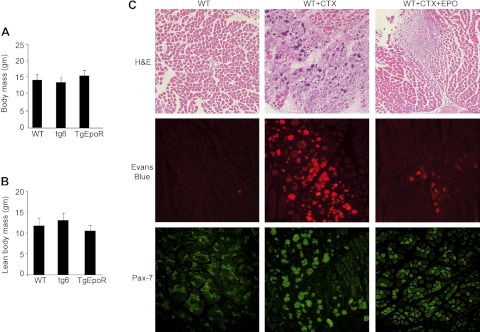
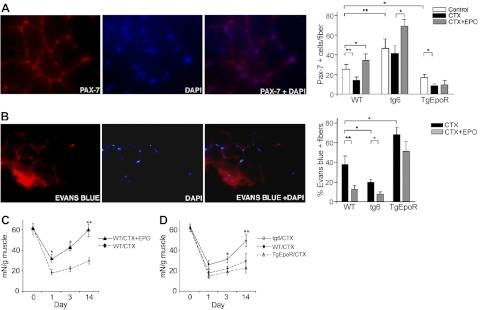
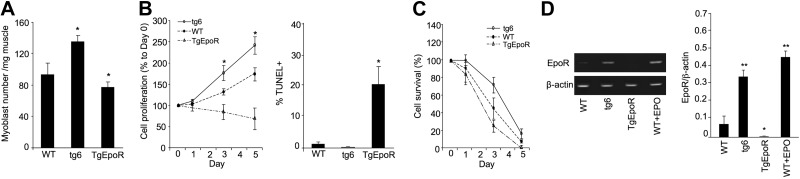
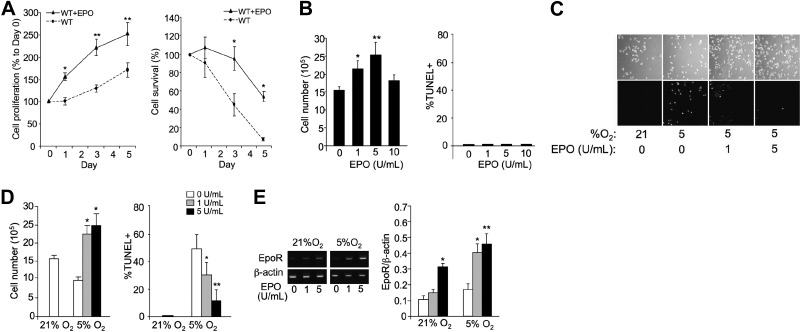
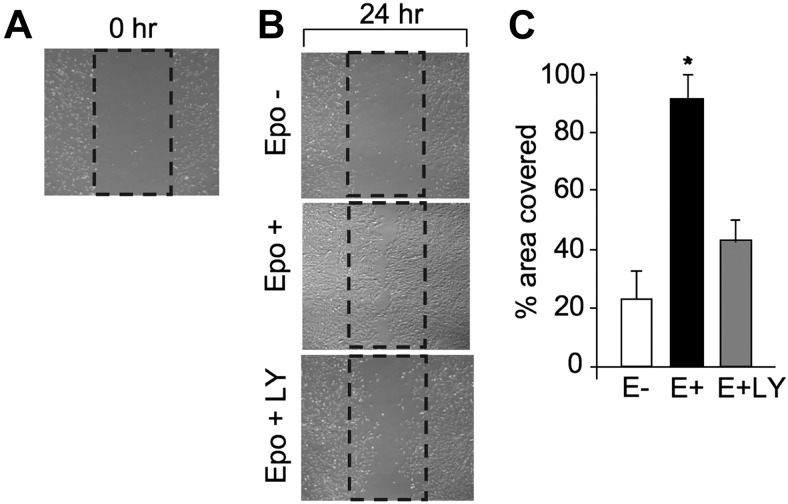
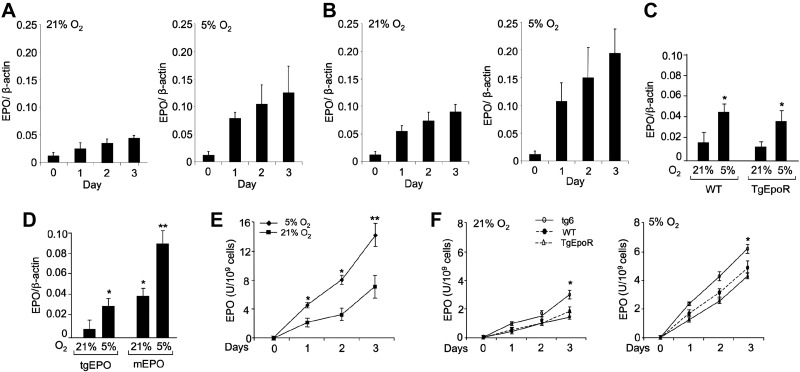
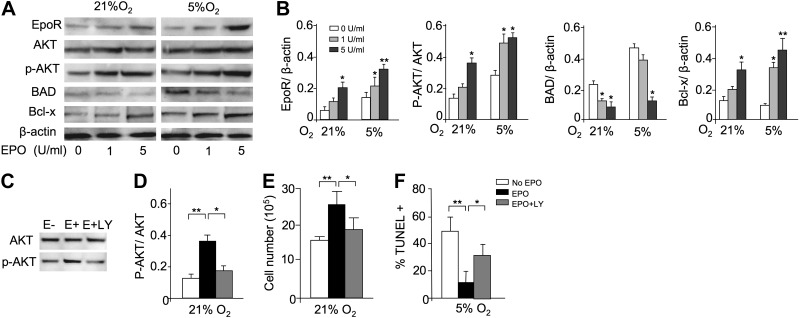
Erythropoietin acts by binding to its cell surface receptor on erythroid progenitor cells to stimulate erythrocyte production. Erythropoietin receptor expression in nonhematopoietic tissue, including skeletal muscle progenitor cells, raises the possibility of a role for erythropoietin beyond erythropoiesis. Mice with erythropoietin receptor restricted to hematopoietic tissue were used to assess contributions of endogenous erythropoietin to promote skeletal myoblast proliferation and survival and wound healing in a mouse model of cardiotoxin induced muscle injury. Compared with wild-type controls, these mice had fewer skeletal muscle Pax-7(+) satellite cells and myoblasts that do not proliferate in culture, were more susceptible to skeletal muscle injury and reduced maximum load tolerated by isolated muscle. In contrast, mice with chronic elevated circulating erythropoietin had more Pax-7(+) satellite cells and myoblasts with increased proliferation and survival in culture, decreased muscle injury, and accelerated recovery of maximum load tolerated by isolated muscle. Skeletal muscle myoblasts also produced endogenous erythropoietin that increased at low O(2). Erythropoietin promoted proliferation, survival, and wound recovery in myoblasts via the phosphoinositide 3-kinase/AKT pathway. Therefore, endogenous and exogenous erythropoietin contribute to increasing satellite cell number following muscle injury, improve myoblast proliferation and survival, and promote repair and regeneration in this mouse induced muscle injury model independent of its effect on erythrocyte production.
Figures

References
-
- Chen Z. Y., Asavaritikrai P., Prchal J. T., Noguchi C. T. (2007) Endogenous erythropoietin signaling is required for normal neural progenitor cell proliferation. J. Biol. Chem. 282, 25875–25883 - PubMed
-
- Ogilvie M., Yu X., Nicolas-Metral V., Pulido S. M., Liu C., Ruegg U. T., Noguchi C. T. (2000) Erythropoietin stimulates proliferation and interferes with differentiation of myoblasts. J. Biol. Chem. 275, 39754–39761 - PubMed
-
- Yu X., Shacka J. J., Eells J. B., Suarez-Quian C., Przygodzki R. M., Beleslin-Cokic B., Lin C. S., Nikodem V. M., Hempstead B., Flanders K. C., Costantini F., Noguchi C. T. (2002) Erythropoietin receptor signalling is required for normal brain development. Development 129, 505–516 - PubMed
-
- Yasuda Y., Masuda S., Chikuma M., Inoue K., Nagao M., Sasaki R. (1998) Estrogen-dependent production of erythropoietin in uterus and its implication in uterine angiogenesis. J. Biol. Chem. 273, 25381–25387 - PubMed
-
- Suzuki N., Ohneda O., Takahashi S., Higuchi M., Mukai H. Y., Nakahata T., Imagawa S., Yamamoto M. (2002) Erythroid-specific expression of the erythropoietin receptor rescued its null mutant mice from lethality. Blood 100, 2279–2288 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
