

PD-1 protects against inflammation and myocyte damage in T cell-mediated myocarditis
- PMID: 22491251
- PMCID: PMC3345066
- DOI: 10.4049/jimmunol.1200389
PD-1 protects against inflammation and myocyte damage in T cell-mediated myocarditis
Abstract
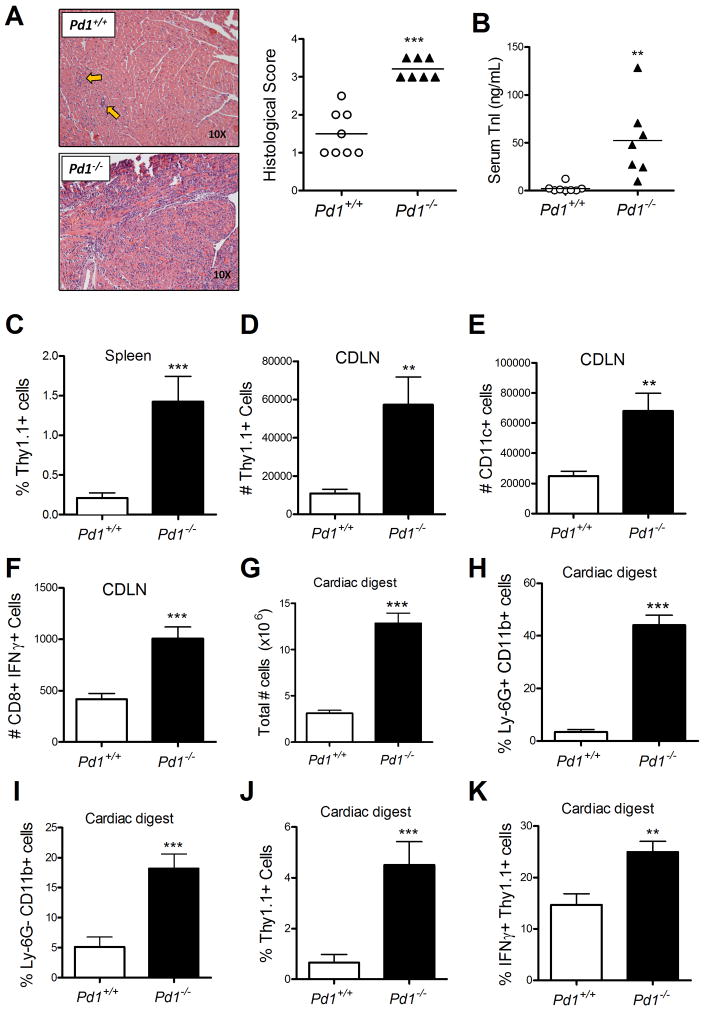
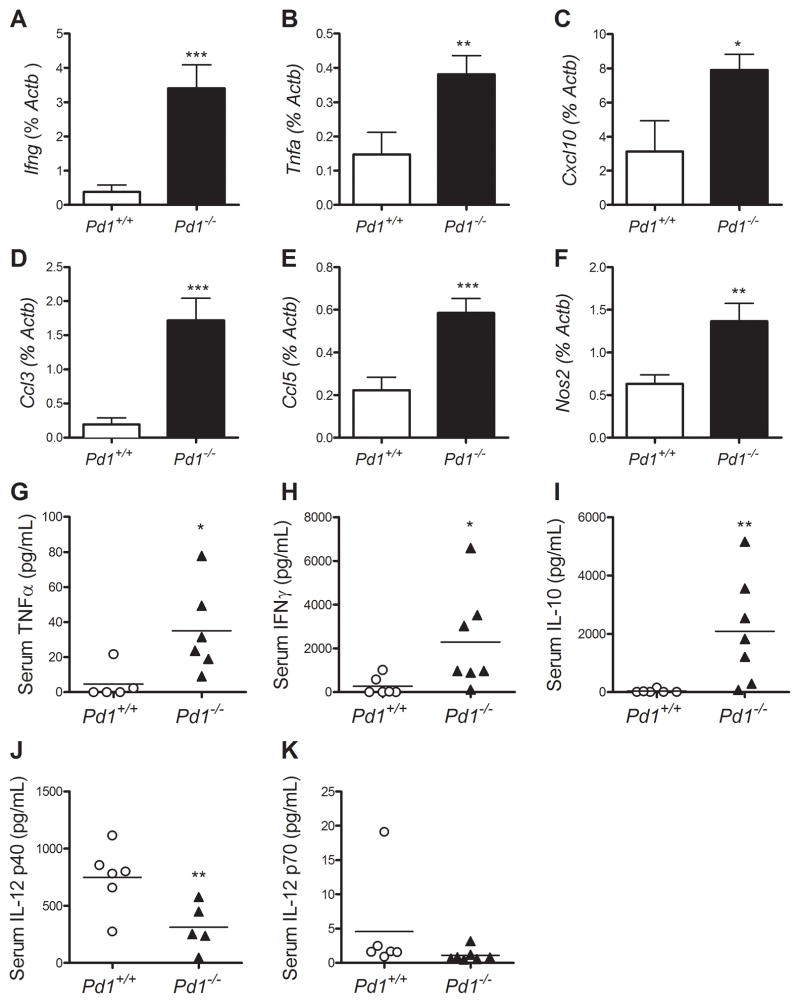
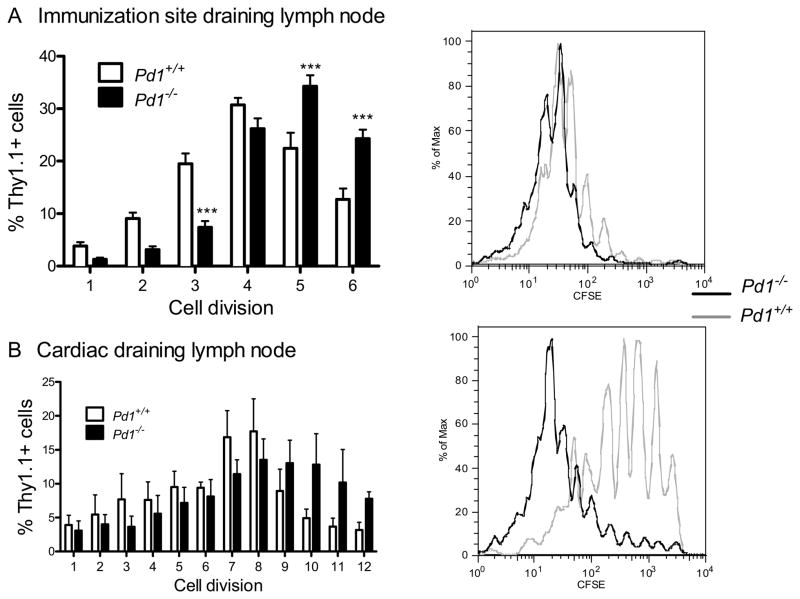
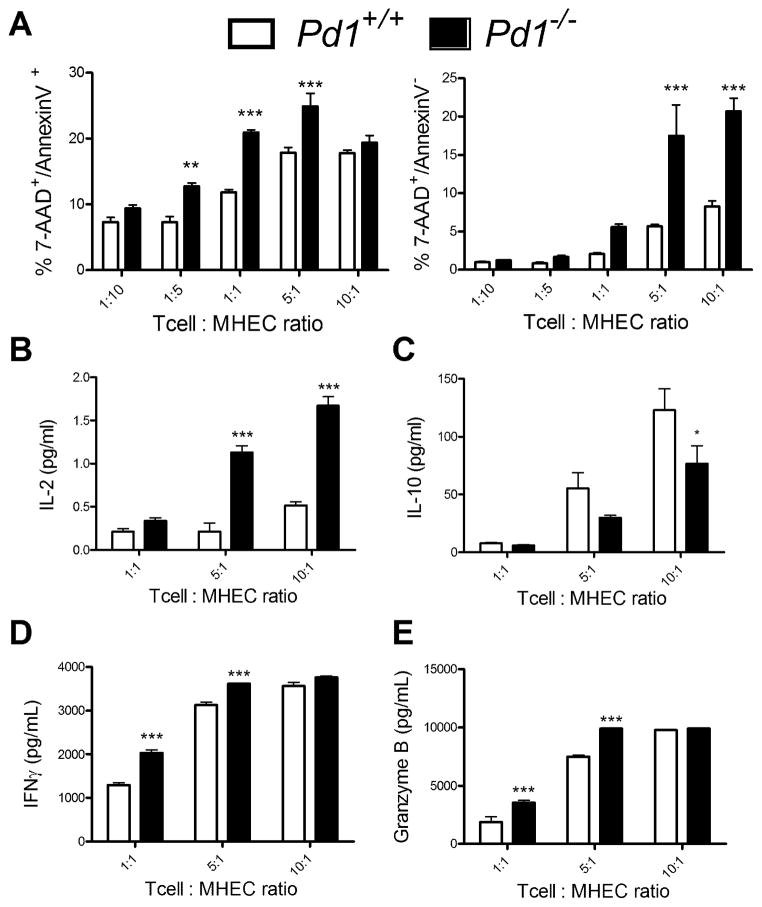
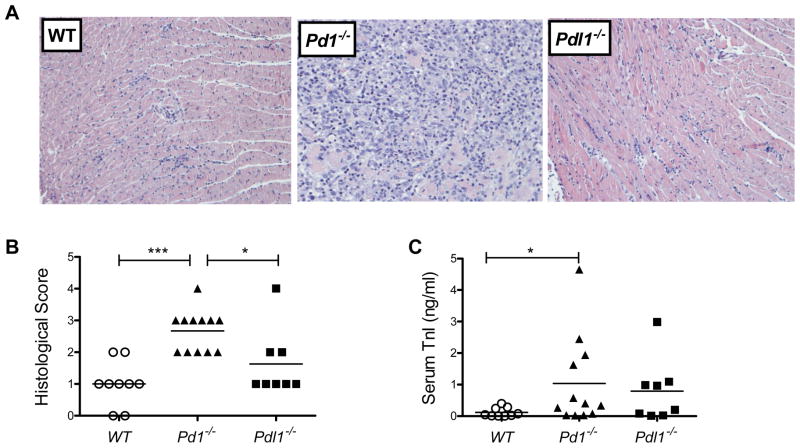
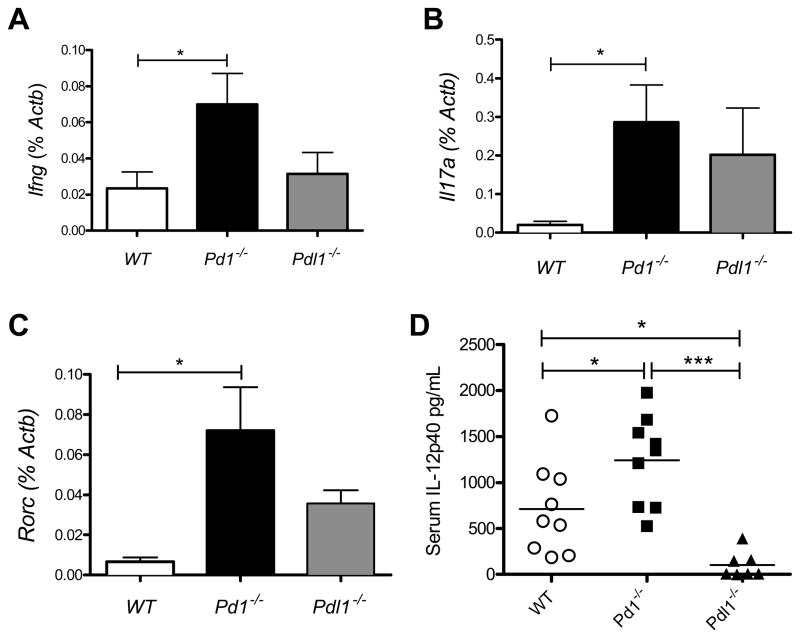
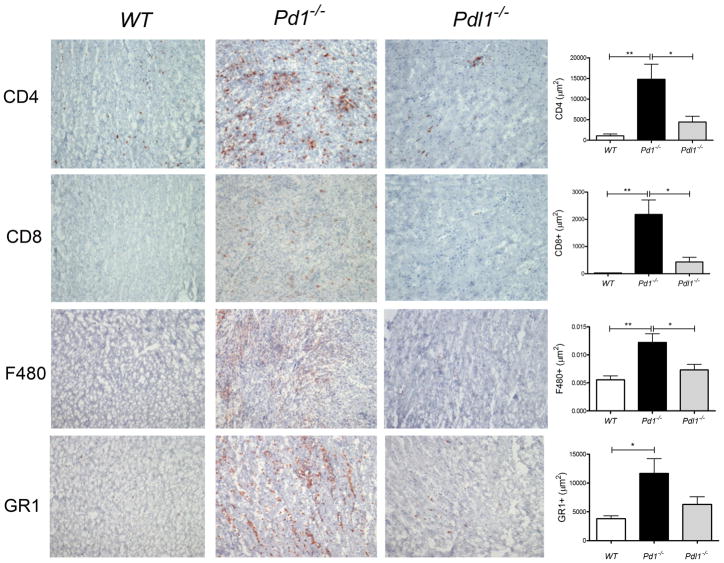
PD-1, a member of the CD28 family of immune regulatory molecules, is expressed on activated T cells, interacts with its ligands, PD-L1/B7-H1 and PD-L2/B7-DC, on other cells, and delivers inhibitory signals to the T cell. We studied the role of this pathway in modulating autoreactive T cell responses in two models of myocarditis. In a CD8(+) T cell-mediated adoptive transfer model, we found that compared with Pd1(+/+) CD8(+) T cells, Pd1(-/-) CD8(+) T cells cause enhanced disease, with increased inflammatory infiltrate, particularly rich in neutrophils. Additionally, we show enhanced proliferation in vivo and enhanced cytotoxic activity of PD-1-deficient T lymphocytes against myocardial endothelial cells in vitro. In experimental autoimmune myocarditis, a disease model dependent on CD4(+) T cells, we show that mice lacking PD-1 develop enhanced disease compared with wild-type mice. PD-1-deficient mice displayed increased inflammation, enhanced serum markers of myocardial damage, and an increased infiltration of inflammatory cells, including CD8(+) T cells. Together, these studies show that PD-1 plays an important role in limiting T cell responses in the heart.
Figures
References
-
- Rose NR, Beisel KW, Herskowitz A, Neu N, Wolfgram LJ, Alvarez FL, Traystman MD, Craig SW. Cardiac myosin and autoimmune myocarditis. Ciba Found Symp. 1987;129:3–24. - PubMed
-
- Cihakova D, Rose NR. Pathogenesis of myocarditis and dilated cardiomyopathy. Adv Immunol. 2008;99:95–114. - PubMed
-
- Baldeviano GC, Barin JG, Talor MV, Srinivasan S, Bedja D, Zheng D, Gabrielson K, Iwakura Y, Rose NR, Cihakova D. Interleukin-17A is dispensable for myocarditis but essential for the progression to dilated cardiomyopathy. Circ Res. 2010;106:1646–1655. - PubMed
-
- Sonderegger I, Rohn TA, Kurrer MO, Iezzi G, Zou Y, Kastelein RA, Bachmann MF, Kopf M. Neutralization of IL-17 by active vaccination inhibits IL-23-dependent autoimmune myocarditis. Eur J Immunol. 2006;36:2849–2856. - PubMed
-
- Afanasyeva M, Georgakopoulos D, Rose NR. Autoimmune myocarditis: cellular mediators of cardiac dysfunction. Autoimmun Rev. 2004;3:476–486. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
