

Complement activation product C5a is a selective suppressor of TLR4-induced, but not TLR3-induced, production of IL-27(p28) from macrophages
- PMID: 22491257
- PMCID: PMC3345104
- DOI: 10.4049/jimmunol.1102914
Complement activation product C5a is a selective suppressor of TLR4-induced, but not TLR3-induced, production of IL-27(p28) from macrophages
Abstract
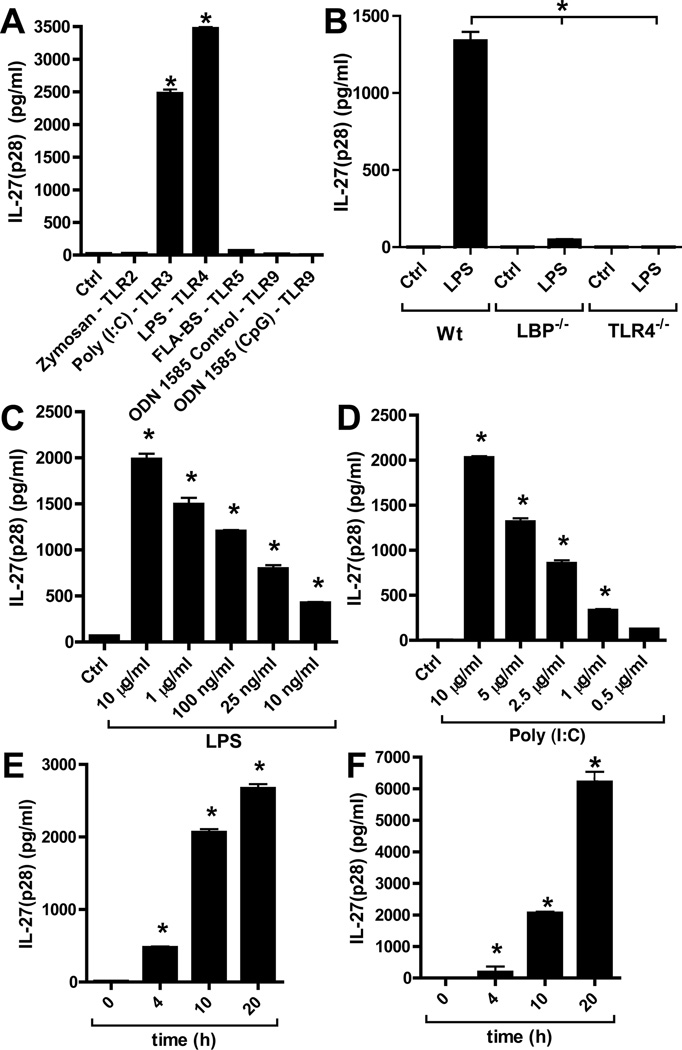
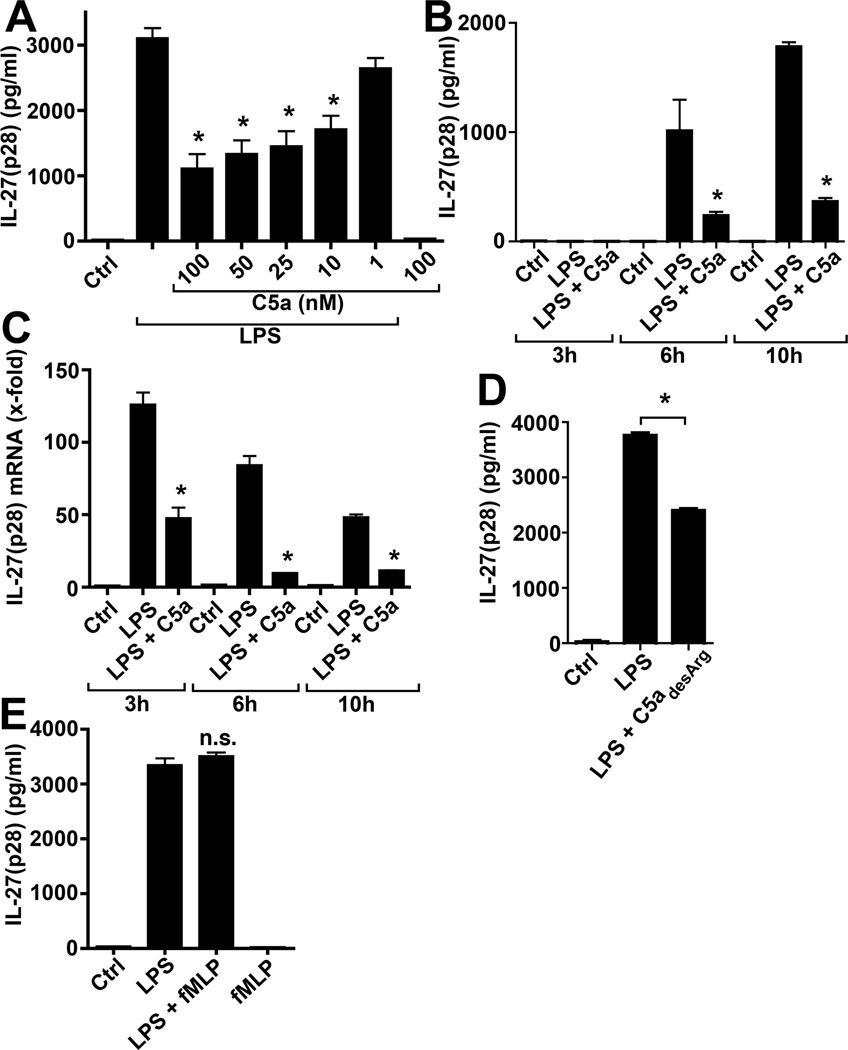
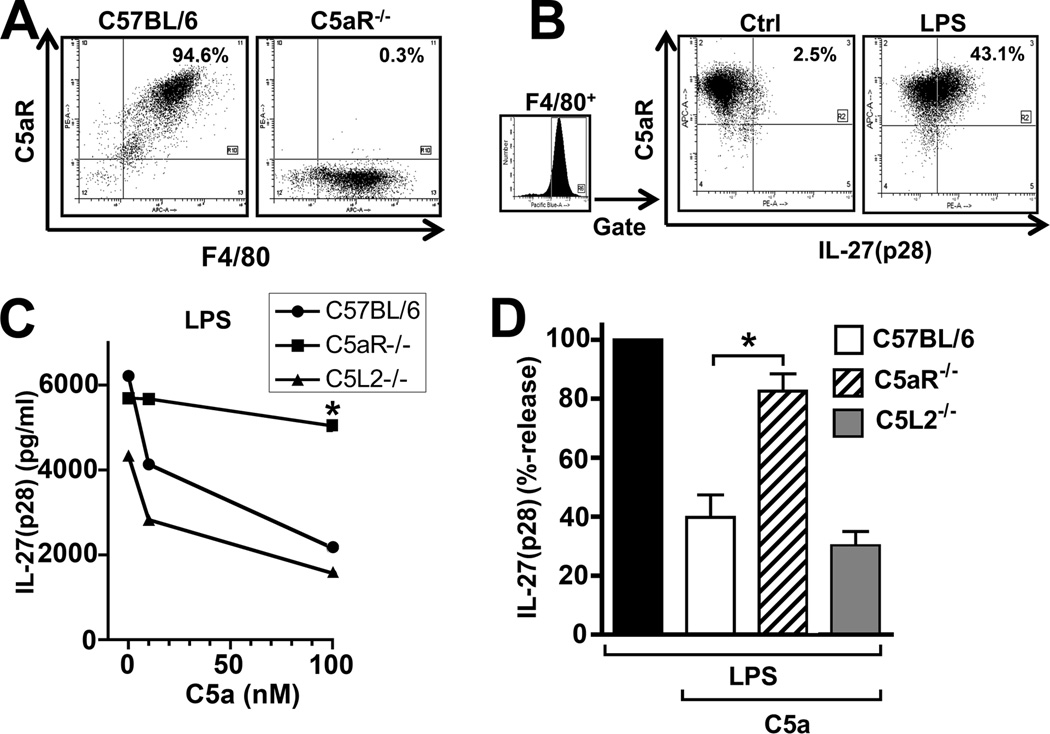
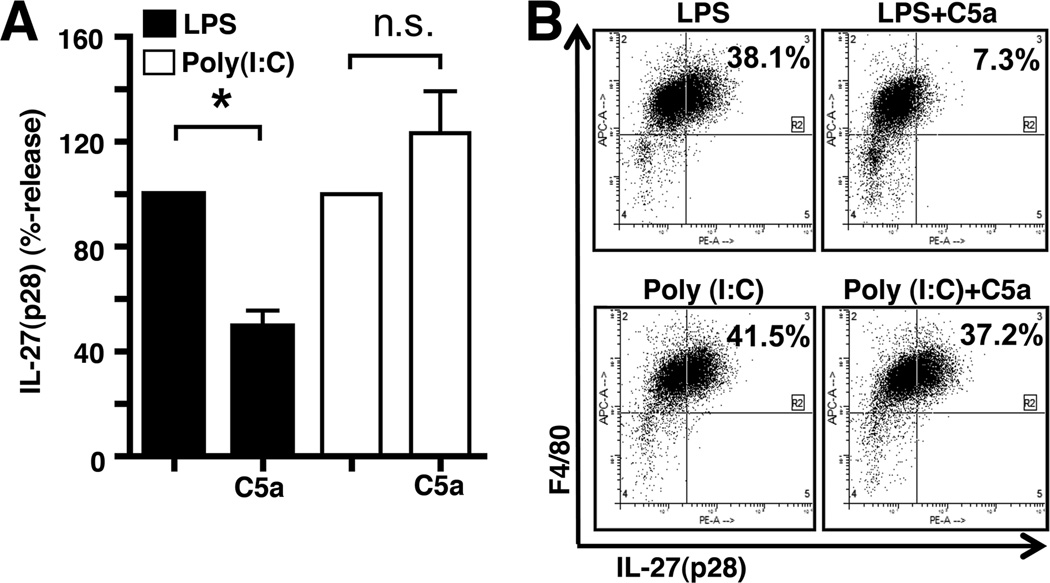
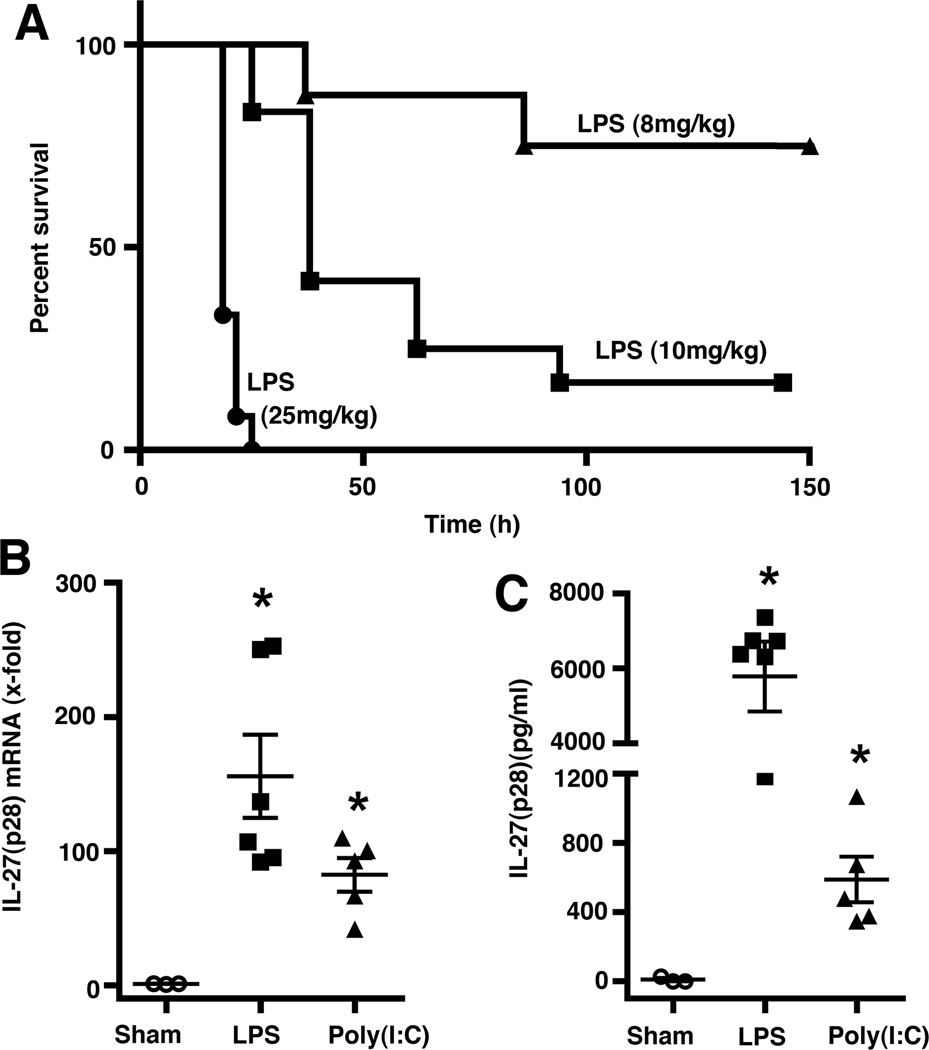
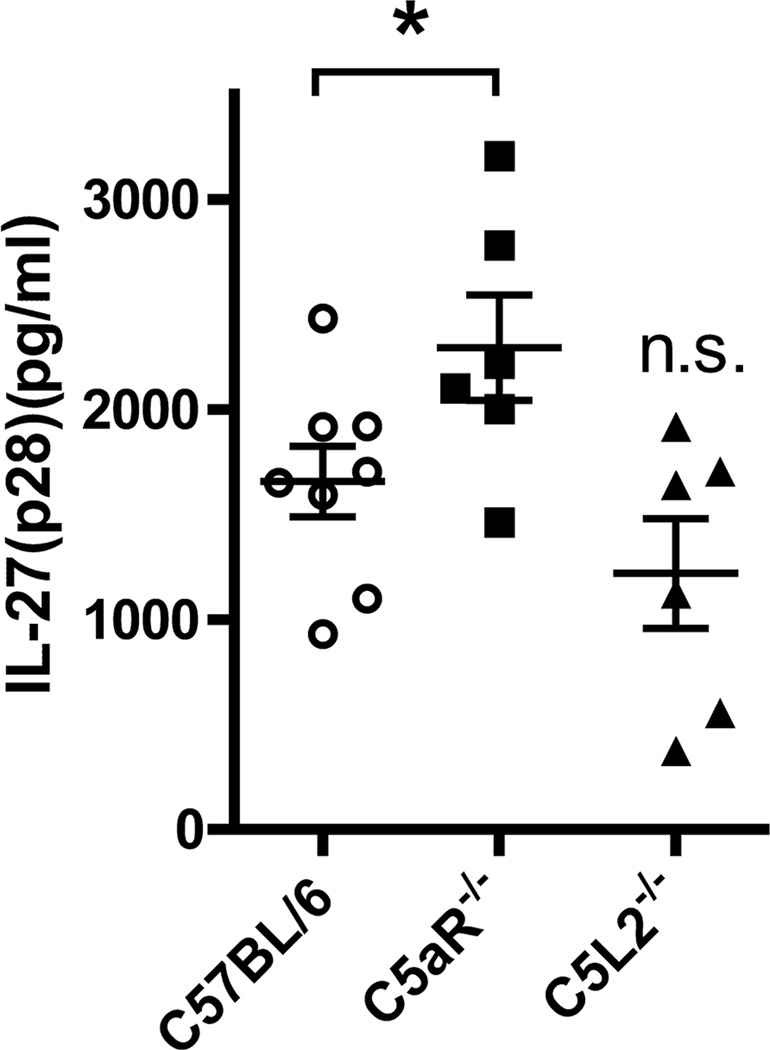
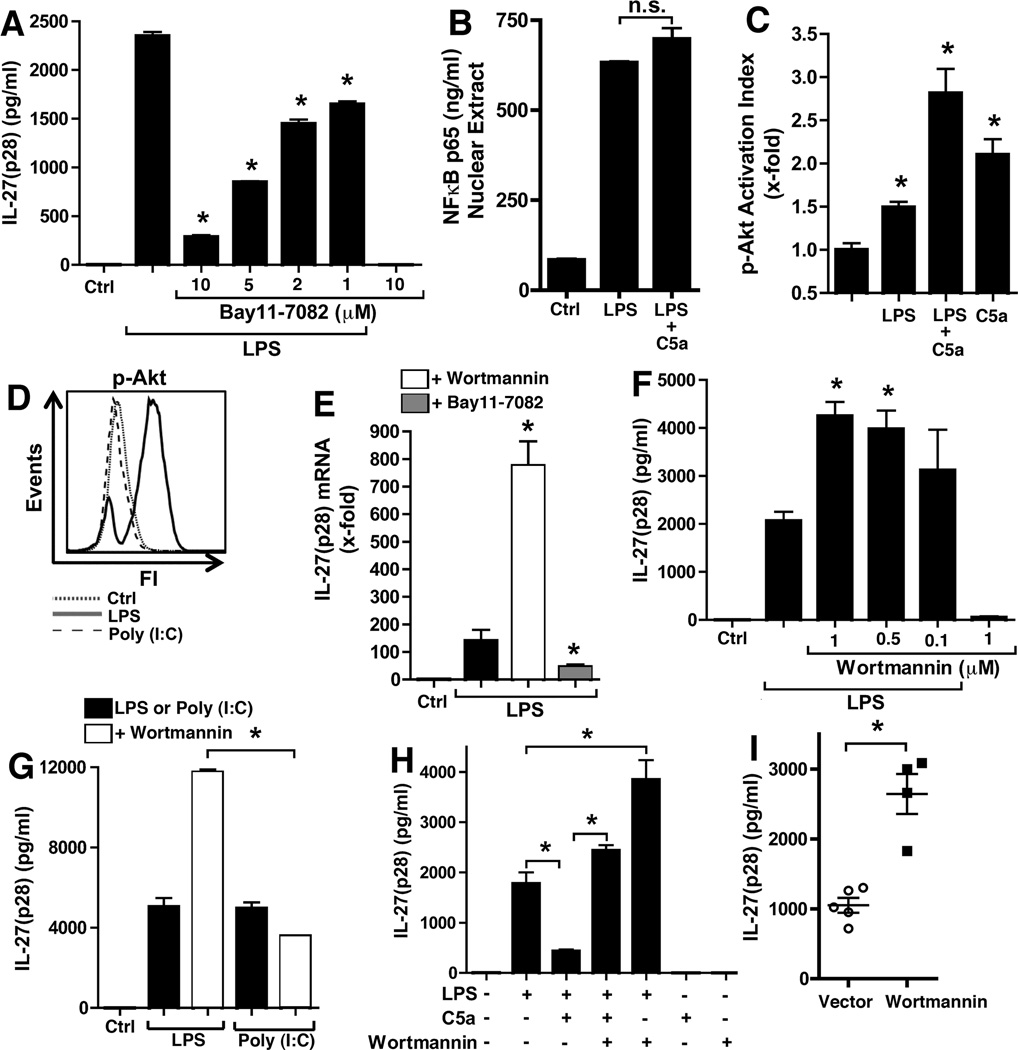
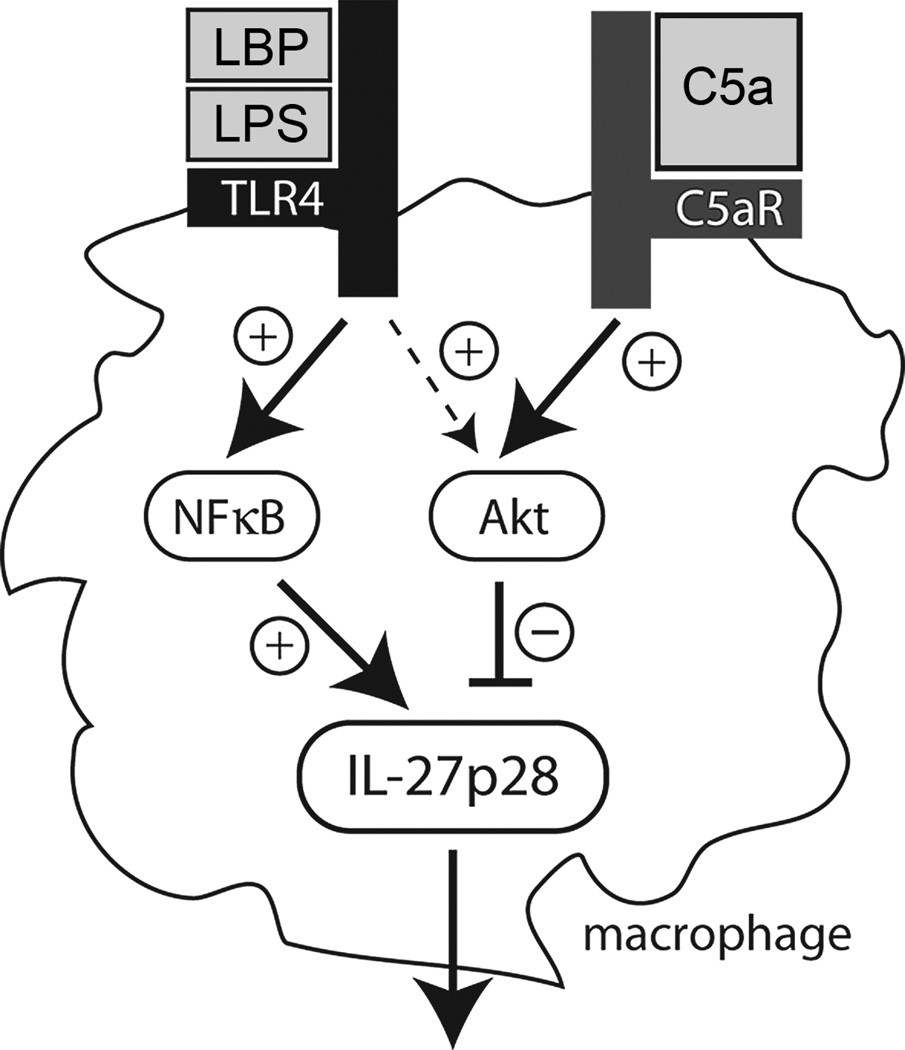
There is accumulating evidence that the complement activation product, C5a, can orchestrate cellular immune functions. IL-27(p28/EBI3) is an emerging key player essential for regulating inflammatory responses and T cells. In this article, we report that C5a robustly suppressed IL-27(p28) gene expression and release in peritoneal macrophages. These cells from C57BL/6J mice abundantly produced IL-27(p28) after engagement of either the TLR3 (polyinosinic-polycytidylic acid) or TLR4 (LPS) receptor. Genetic deficiency of either TLR4 or LBP completely incapacitated the ability of macrophages to secrete IL-27(p28) in response to LPS. IL-27(p28)-producing macrophages also expressed the C5aR receptor, thus displaying an IL-27(p28)(+)F4/80(+)C5aR(+) phenotype. C5a suppressed IL-27(p28) in LPS-stimulated macrophages via interactions with the C5aR receptor rather than the C5L2 receptor. After endotoxemia, C5aR(-/-) mice displayed higher plasma levels of IL-27(p28) compared with C57BL/6J mice. C5a did not affect the release of IL-27(p28) or the frequency of IL-27(p28)(+)F4/80(+) macrophages after engagement of TLR3. Mechanistically, LPS activated both the NF-κB and the PI3K/Akt pathways, whereas C5a activated only the PI3K/Akt pathway. Engagement of PI3K/Akt was inhibitory for IL-27(p28) production, because PI3K/Akt pharmacologic blockade resulted in increased amounts of IL-27(p28) and reversed the suppressive effects of C5a. Blockade of PI3K/Akt in endotoxemic C57BL/6J mice resulted in higher generation of IL-27(p28). In contrast, the PI3K/Akt pathway was not involved in TLR3-mediated release of IL-27(p28). These data provide new evidence about how complement activation may selectively interfere with production of T cell regulatory cytokines by APCs in the varying contexts of either bacterial (TLR4 pathway) or viral (TLR3 pathway) infection.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
