

Neural mechanisms of foraging
- PMID: 22491854
- PMCID: PMC3440844
- DOI: 10.1126/science.1216930
Neural mechanisms of foraging
Abstract
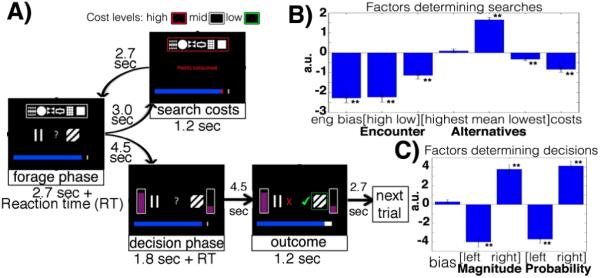
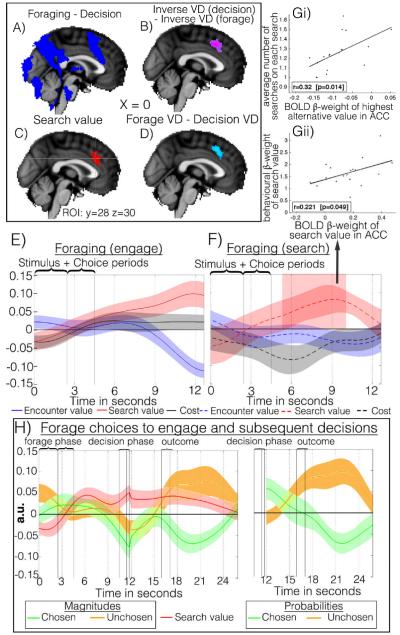
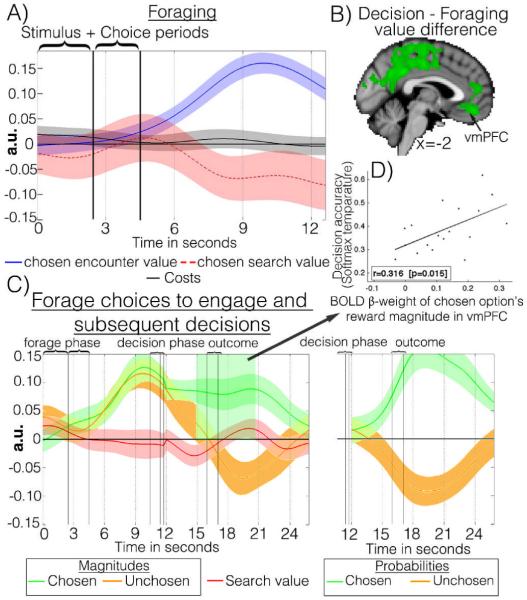
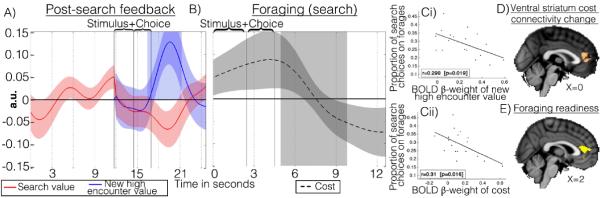
Behavioral economic studies involving limited numbers of choices have provided key insights into neural decision-making mechanisms. By contrast, animals' foraging choices arise in the context of sequences of encounters with prey or food. On each encounter, the animal chooses whether to engage or, if the environment is sufficiently rich, to search elsewhere. The cost of foraging is also critical. We demonstrate that humans can alternate between two modes of choice, comparative decision-making and foraging, depending on distinct neural mechanisms in ventromedial prefrontal cortex (vmPFC) and anterior cingulate cortex (ACC) using distinct reference frames; in ACC, choice variables are represented in invariant reference to foraging or searching for alternatives. Whereas vmPFC encodes values of specific well-defined options, ACC encodes the average value of the foraging environment and cost of foraging.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
