

Transcriptional co-regulation of neuronal migration and laminar identity in the neocortex
- PMID: 22492350
- PMCID: PMC3317962
- DOI: 10.1242/dev.069963
Transcriptional co-regulation of neuronal migration and laminar identity in the neocortex
Abstract
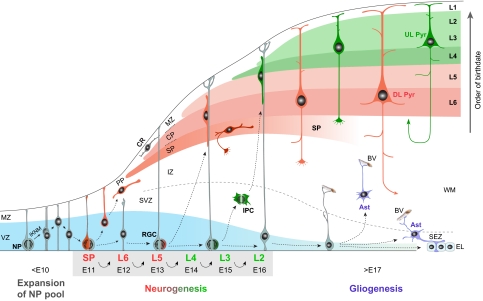
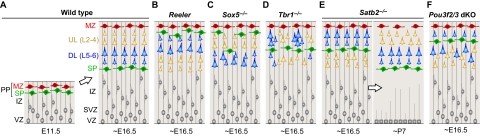
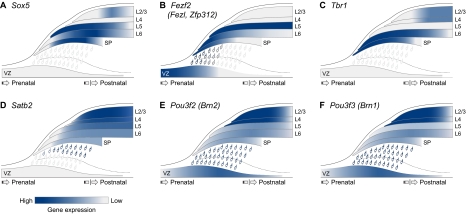
The cerebral neocortex is segregated into six horizontal layers, each containing unique populations of molecularly and functionally distinct excitatory projection (pyramidal) neurons and inhibitory interneurons. Development of the neocortex requires the orchestrated execution of a series of crucial processes, including the migration of young neurons into appropriate positions within the nascent neocortex, and the acquisition of layer-specific neuronal identities and axonal projections. Here, we discuss emerging evidence supporting the notion that the migration and final laminar positioning of cortical neurons are also co-regulated by cell type- and layer-specific transcription factors that play concomitant roles in determining the molecular identity and axonal connectivity of these neurons. These transcriptional programs thus provide direct links between the mechanisms controlling the laminar position and identity of cortical neurons.
Figures

References
-
- Akbarian S., Kim J. J., Potkin S. G., Hetrick W. P., Bunney W. E., Jones E. G. (1996). Maldistribution of interstitial neurons in prefrontal white matter of the brains of schizophrenic patients. Arch. Gen. Psychiatry 53, 425–436 - PubMed
-
- Alcamo E. A., Chirivella L, Dautzenberg M, Dobreva G, Fariñas I, Grosschedl R., McConnell S. K. (2008). Satb2 regulates callosal projection neuron identity in the developing cerebral cortex. Neuron 57, 364–377 - PubMed
-
- Alfano C., Viola L., Heng J. I., Pirozzi M., Clarkson M., Flore G., De Maio A., Schedl A., Guillemot F., Studer M. (2011). COUP-TFI promotes radial migration and proper morphology of callosal projection neurons by repressing Rnd2 expression. Development 138, 4685–4697 - PubMed
-
- Allendoerfer K. L., Shatz C. J. (1994). The subplate, a transient neocortical structure: its role in the development of connections between thalamus and cortex. Annu. Rev. Neurosci. 17, 185–218 - PubMed
-
- Anderson S. A., Volk D. W., Lewis D. A. (1996). Increased density of microtubule associated protein 2-immunoreactive neurons in the prefrontal white matter of schizophrenic subjects. Schizophr. Res. 19, 111–119 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
