

Dynamic in vivo binding of transcription factors to cis-regulatory modules of cer and gsc in the stepwise formation of the Spemann-Mangold organizer
- PMID: 22492356
- PMCID: PMC4074222
- DOI: 10.1242/dev.068395
Dynamic in vivo binding of transcription factors to cis-regulatory modules of cer and gsc in the stepwise formation of the Spemann-Mangold organizer
Abstract
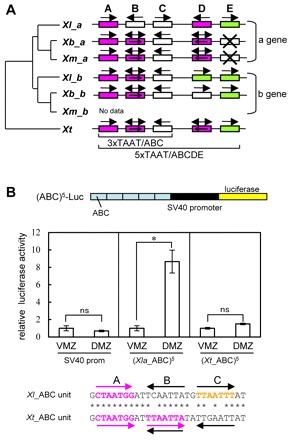
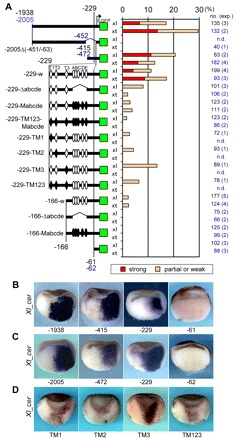
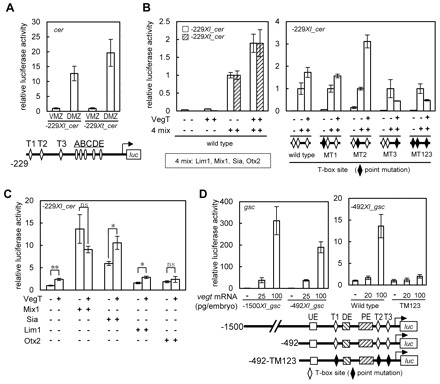
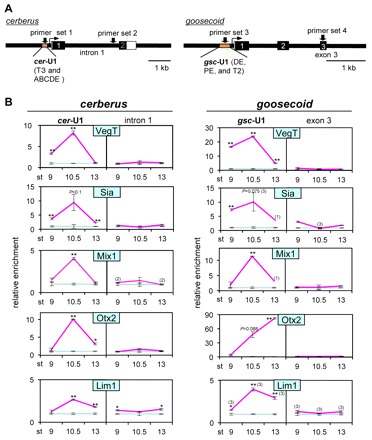
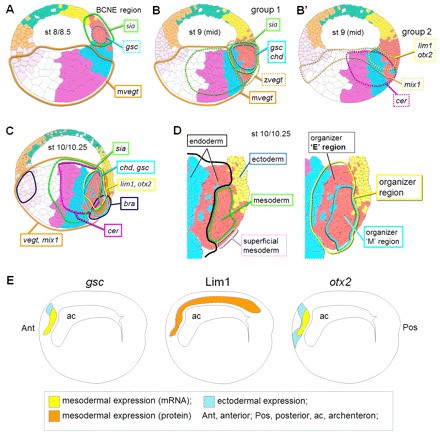
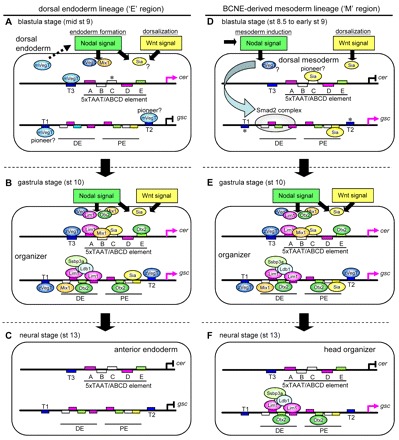
How multiple developmental cues are integrated on cis-regulatory modules (CRMs) for cell fate decisions remains uncertain. The Spemann-Mangold organizer in Xenopus embryos expresses the transcription factors Lim1/Lhx1, Otx2, Mix1, Siamois (Sia) and VegT. Reporter analyses using sperm nuclear transplantation and DNA injection showed that cerberus (cer) and goosecoid (gsc) are activated by the aforementioned transcription factors through CRMs conserved between X. laevis and X. tropicalis. ChIP-qPCR analysis for the five transcription factors revealed that cer and gsc CRMs are initially bound by both Sia and VegT at the late blastula stage, and subsequently bound by all five factors at the gastrula stage. At the neurula stage, only binding of Lim1 and Otx2 to the gsc CRM, among others, persists, which corresponds to their co-expression in the prechordal plate. Based on these data, together with detailed expression pattern analysis, we propose a new model of stepwise formation of the organizer, in which (1) maternal VegT and Wnt-induced Sia first bind to CRMs at the blastula stage; then (2) Nodal-inducible Lim1, Otx2, Mix1 and zygotic VegT are bound to CRMs in the dorsal endodermal and mesodermal regions where all these genes are co-expressed; and (3) these two regions are combined at the gastrula stage to form the organizer. Thus, the in vivo dynamics of multiple transcription factors highlight their roles in the initiation and maintenance of gene expression, and also reveal the stepwise integration of maternal, Nodal and Wnt signaling on CRMs of organizer genes to generate the organizer.
Figures
References
-
- Bauer D. V., Huang S., Moody S. A. (1994). The cleavage stage origin of Spemann’s Organizer: analysis of the movements of blastomere clones before and during gastrulation in Xenopus. Development 120, 1179–1189 - PubMed
-
- Blitz I. L., Cho K. W. (1995). Anterior neurectoderm is progressively induced during gastrulation: the role of the Xenopus homeobox gene orthodenticle. Development 121, 993–1004 - PubMed
-
- Bouwmeester T., Kim S., Sasai Y., Lu B., De Robertis E. M. (1996). Cerberus is a head-inducing secreted factor expressed in the anterior endoderm of Spemann’s organizer. Nature 382, 595–601 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
