

Analysis of centriole elimination during C. elegans oogenesis
- PMID: 22492357
- PMCID: PMC4074223
- DOI: 10.1242/dev.075440
Analysis of centriole elimination during C. elegans oogenesis
Abstract
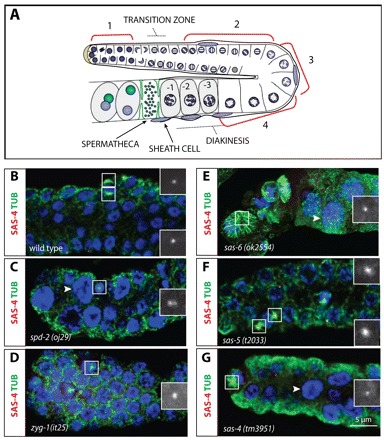
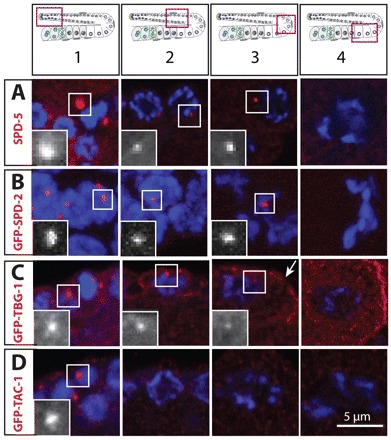
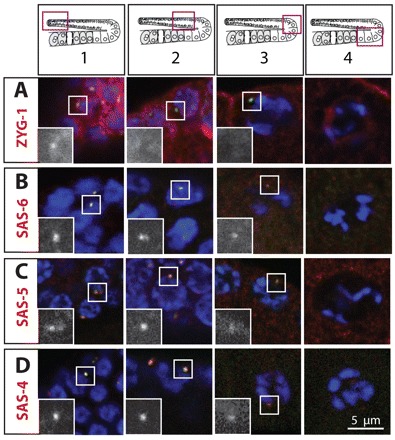
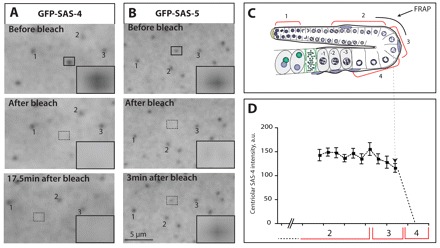
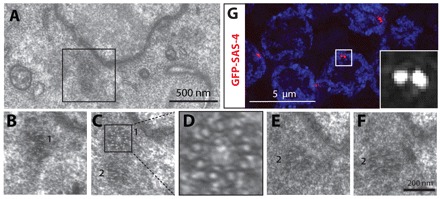
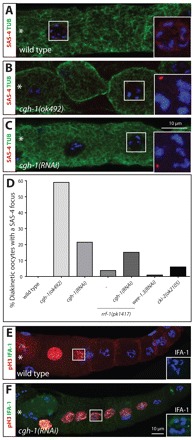
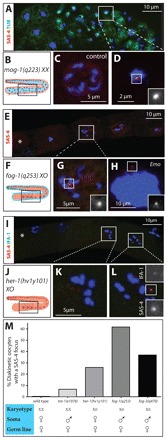
Centrosomes are the principal microtubule organizing centers (MTOCs) of animal cells and comprise a pair of centrioles surrounded by pericentriolar material (PCM). Centriole number must be carefully regulated, notably to ensure bipolar spindle formation and thus faithful chromosome segregation. In the germ line of most metazoan species, centrioles are maintained during spermatogenesis, but eliminated during oogenesis. Such differential behavior ensures that the appropriate number of centrioles is present in the newly fertilized zygote. Despite being a fundamental feature of sexual reproduction in metazoans, the mechanisms governing centriole elimination during oogenesis are poorly understood. Here, we investigate this question in C. elegans. Using antibodies directed against centriolar components and serial-section electron microscopy, we establish that centrioles are eliminated during the diplotene stage of the meiotic cell cycle. Moreover, we show that centriole elimination is delayed upon depletion of the helicase CGH-1. We also find that somatic cells make a minor contribution to this process, and demonstrate that the germ cell karyotype is important for timely centriole elimination. These findings set the stage for a mechanistic dissection of centriole elimination in a metazoan organism.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
